
Homo erectus: Difference between revisions
m Homo erectus |
Dudley Miles (talk | contribs) Deleted cladograms not supported by sources |
||
| Line 1: | Line 1: | ||
{{short description|Extinct species of archaic human}} |
|||
{{redirect|H. erectus|the seahorse species|Hippocampus erectus|the 2007 comedy film|Homo Erectus (film)|1997 album|Homo erectus (album)}} |
|||
{{redirect| |
{{redirect|H. erectus|other uses|H. erectus (disambiguation)|and|Homo erectus (disambiguation)}} |
||
{{redirect|Pithecanthropus erectus|the song and album by Charles Mingus|Pithecanthropus Erectus (album)}} |
|||
{{pp-move-indef}} |
|||
{{pp-move}} |
|||
{{Use dmy dates|date=July 2013}} |
|||
{{Use dmy dates|date=September 2020}} |
|||
{{italic title}} |
|||
{{Speciesbox |
|||
{{Taxobox |
|||
| fossil_range = {{Fossil range|2|0.1}}<small>[[Early Pleistocene]] – [[Late Pleistocene]]<ref name=Rizal/></small> |
|||
| name = ''Homo erectus'' |
|||
| image = Peking Man Skull (replica) presented at Paleozoological Museum of China.jpg |
|||
| fossil_range = {{Fossil range|1.9|0.07}}<small>Early [[Pleistocene]] – Middle [[Pleistocene]]</small> |
|||
| image_caption = Replica of the skull of [[Peking Man]] at the [[Paleozoological Museum of China]] |
|||
| image = Homme de Tautavel 01-08.jpg |
|||
| |
| extinct = yes |
||
| taxon = Homo erectus |
|||
| image_caption = Reconstruction of a specimen from Tautavel, France |
|||
| authority = ([[Eugène Dubois|Dubois]], 1893) |
|||
| regnum = [[Animal]]ia |
|||
| synonyms = * ''[[Java Man|Anthropopithecus erectus]]'' <small>[[Eugène Dubois|Dubois]], 1893</small> |
|||
| phylum = [[Chordate|Chordata]] |
|||
* ''[[Java Man|Pithecanthropus erectus]]'' <small>([[Eugène Dubois|Dubois]], 1893)</small> |
|||
| classis = [[Mammal]]ia |
|||
* ''[[Peking Man|Sinanthropus pekinensis]]'' |
|||
| ordo = [[Primates]] |
|||
* ''[[Solo Man|Javanthropus soloensis]]'' |
|||
| familia = [[Hominid]]ae |
|||
* ''[[Tighennif|Atlanthropus mauritanicus]]'' |
|||
| genus = ''[[Homo (genus)|Homo]]'' |
|||
* (?) ''[[Homo ergaster|Telanthropus capensis]]'' |
|||
| species = '''''H. erectus''''' |
|||
* (?) ''[[Homo georgicus]]'' |
|||
* (?) ''[[Tautavel Man|Homo tautavelensis]]'' |
|||
| binomial_authority = ([[Eugène Dubois|Dubois]], 1892) |
|||
}} |
|||
| synonyms = |
|||
*† ''[[Java Man|Anthropopithecus erectus]]'' |
|||
*† ''[[Java Man|Pithecanthropus erectus]]'' |
|||
*† ''[[Sinanthropus pekinensis]]'' |
|||
*† ''Javanthropus soloensis'' |
|||
*† ''Meganthropus paleojavanicus'' |
|||
*† ''Telanthropus capensis'' |
|||
*† ''Homo georgicus'' |
|||
*† ''[[Homo ergaster]]''? |
|||
}} |
|||
'''''Homo erectus ''''' (meaning "upright man", from the Latin ''ērigere'', "to put up, set upright") is an extinct [[species]] of [[hominid]] that lived throughout most of the [[Pleistocene]] geological epoch. Its earliest fossil evidence dates to 1.9 million years ago and the most recent to 70,000 years ago. Its extinction is linked by some scientists to the [[Toba catastrophe theory|Toba super-eruption catastrophe]], but no sufficient case has been made to date for the idea. It is generally thought that ''H. erectus'' originated in [[Africa]] and spread from there, migrating throughout Eurasia as far as [[Georgia (country)|Georgia]], [[India]], [[Sri Lanka]], [[China]] and [[Java]].<ref name="Hazarika">{{cite news|last=Hazarika|first=Manji|title=''Homo erectus/ergaster'' and Out of Africa: Recent Developments in Paleoanthropology and Prehistoric Archaeology|date=16–30 June 2007|url=http://www.himalayanlanguages.org/files/hazarika/Manjil%20Hazarika%20EAA.pdf}}</ref><ref name="Chauhan">Chauhan, Parth R. (2003) [http://www.assemblage.group.shef.ac.uk/issue7/chauhan.html#distribution "Distribution of Acheulian sites in the Siwalik region"] in ''An Overview of the Siwalik Acheulian & Reconsidering Its Chronological Relationship with the Soanian – A Theoretical Perspective''. assemblage.group.shef.ac.uk</ref> But other scientists posit that the species rose first, or separately, in Asia. |
|||
'''''Homo erectus''''' ({{IPAc-en|ˌ|h|oʊ|m|oʊ|_|ə|'|r|ɛ|k|t|ə|s}} {{lit|[[:wikt:erectus|upright]] man}}) is an [[Extinction|extinct]] [[species]] of [[archaic human]] from the [[Pleistocene]], with its earliest occurrence about 2 million years ago.<ref name=Herries>{{cite journal | vauthors = Herries AI, Martin JM, Leece AB, Adams JW, Boschian G, Joannes-Boyau R, Edwards TR, Mallett T, Massey J, Murszewski A, Neubauer S, Pickering R, Strait DS, Armstrong BJ, Baker S, Caruana MV, Denham T, Hellstrom J, Moggi-Cecchi J, Mokobane S, Penzo-Kajewski P, Rovinsky DS, Schwartz GT, Stammers RC, Wilson C, Woodhead J, Menter C | display-authors = 6 | title = Contemporaneity of ''Australopithecus'', ''Paranthropus'', and early ''Homo erectus'' in South Africa | journal = Science | volume = 368 | issue = 6486 | pages = eaaw7293 | date = April 2020 | pmid = 32241925 | doi = 10.1126/science.aaw7293 | doi-access = free }}</ref> Its specimens are among the first recognizable members of the genus ''[[Homo]]''. |
|||
Debate also continues about the classification, ancestry, and progeny of ''Homo erectus'', especially vis-à-vis ''[[Homo ergaster]]'', with two major positions: 1) ''H. erectus'' is the same species as ''[[H. ergaster]]'', and thereby ''H. erectus'' is a direct ancestor of the later hominins including ''[[Homo heidelbergensis]]'', ''[[Homo neanderthalensis]]'', and ''[[Homo sapiens]]''; or, 2) it is in fact an [[Asia]]n species distinct from African ''H. ergaster''.<ref name="Hazarika"/><ref>See overview of theories on [[human evolution]].</ref><ref>Klein, R. (1999). ''The Human Career: Human Biological and Cultural Origins''. Chicago: University of Chicago Press, ISBN 0226439631.</ref> |
|||
Several human species, such as ''[[H. heidelbergensis]]'' and ''[[H. antecessor]]'', appear to have evolved from ''H. erectus'', and [[Neanderthal]]s, [[Denisovan]]s, and [[modern humans]] are in turn generally considered to have evolved from ''H. heidelbergensis''.<ref>{{cite journal | vauthors = Dembo M, Radovčić D, Garvin HM, Laird MF, Schroeder L, Scott JE, Brophy J, Ackermann RR, Musiba CM, de Ruiter DJ, Mooers AØ, Collard M | display-authors = 6 | title = The evolutionary relationships and age of Homo naledi: An assessment using dated Bayesian phylogenetic methods | journal = Journal of Human Evolution | volume = 97 | pages = 17–26 | date = August 2016 | pmid = 27457542 | doi = 10.1016/j.jhevol.2016.04.008 | bibcode = 2016JHumE..97...17D | hdl = 2164/8796 | hdl-access = free }}</ref> ''H. erectus'' was the first human ancestor to spread throughout [[Eurasia]], with a continental range extending from the [[Iberian Peninsula]] to [[Java]]. Asian populations of ''H. erectus'' may be ancestral to ''[[H. floresiensis]]''<ref>{{cite journal | vauthors = van den Bergh GD, Kaifu Y, Kurniawan I, Kono RT, Brumm A, Setiyabudi E, Aziz F, Morwood MJ | display-authors = 6 | title = Homo floresiensis-like fossils from the early Middle Pleistocene of Flores | journal = Nature | volume = 534 | issue = 7606 | pages = 245–248 | date = June 2016 | pmid = 27279221 | doi = 10.1038/nature17999 | bibcode = 2016Natur.534..245V | s2cid = 205249218 | author5-link = Adam Brumm }}</ref> and possibly to ''[[Homo luzonensis|H. luzonensis]]''.<ref>{{cite journal | vauthors = Détroit F, Mijares AS, Corny J, Daver G, Zanolli C, Dizon E, Robles E, Grün R, Piper PJ | display-authors = 6 | title = A new species of Homo from the Late Pleistocene of the Philippines | journal = Nature | volume = 568 | issue = 7751 | pages = 181–186 | date = April 2019 | pmid = 30971845 | doi = 10.1038/s41586-019-1067-9 | s2cid = 106411053 | bibcode = 2019Natur.568..181D | url = https://hal.archives-ouvertes.fr/hal-02296712/file/Detroit_%26_al_2019_Nature_postprint.pdf }}</ref> The last known population of ''H. erectus'' is ''[[Solo Man|H. e. soloensis]]'' from Java, around 117,000–108,000 years ago.<ref name=Rizal>{{cite journal | vauthors = Rizal Y, Westaway KE, Zaim Y, van den Bergh GD, Bettis EA, Morwood MJ, Huffman OF, Grün R, Joannes-Boyau R, Bailey RM, Westaway MC, Kurniawan I, Moore MW, Storey M, Aziz F, Zhao JX, Sipola ME, Larick R, Zonneveld JP, Scott R, Putt S, Ciochon RL | display-authors = 6 | title = Last appearance of Homo erectus at Ngandong, Java, 117,000-108,000 years ago | journal = Nature | volume = 577 | issue = 7790 | pages = 381–385 | date = January 2020 | pmid = 31853068 | doi = 10.1038/s41586-019-1863-2 | s2cid = 209410644 }}</ref> |
|||
And there is another view—an alternative to 1): some [[palaeoanthropologist]]s consider ''H. ergaster'' to be a variety, that is, the "''African''" variety, of ''H. erectus'', and they offer the labels "''Homo erectus [[sensu stricto]]''" (strict sense) for the Asian species and "''Homo erectus [[sensu lato]]''" (broad sense) for the greater species comprising both Asian and African populations.<!------------><ref>{{cite journal|author=Antón, S. C. |year=2003|title=Natural history of Homo erectus|journal= Am. J. Phys. Anthropol.|volume=122|pages=126–170|doi=10.1002/ajpa.10399|quote=By the 1980s, the growing numbers of ''H. erectus'' specimens, particularly in Africa, led to the realization that Asian ''H. erectus'' (''H. erectus sensu stricto''), once thought so primitive, was in fact more derived than its African counterparts. These morphological differences were interpreted by some as evidence that more than one species might be included in ''H. erectus sensu lato'' (e.g., Stringer, 1984; Andrews, 1984; Tattersall, 1986; Wood, 1984, 1991a, b; Schwartz and Tattersall, 2000) ... Unlike the European lineage, in my opinion, the taxonomic issues surrounding Asian vs. African H. erectus are more intractable. The issue was most pointedly addressed with the naming of H. ergaster on the basis of the type mandible KNM-ER 992, but also including the partial skeleton and isolated teeth of KNM-ER 803 among other Koobi Fora remains (Groves and Mazak, 1975). Recently, this specific name was applied to most early African and Georgian H. erectus in recognition of the less-derived nature of these remains vis à vis conditions in Asian H. erectus (see Wood, 1991a, p. 268; Gabunia et al., 2000a). It should be noted, however, that at least portions of the paratype of H. ergaster (e.g., KNM-ER 1805) are not included in most current conceptions of that taxon. The ''H. ergaster'' question remains famously unresolved (e.g., Stringer, 1984; Tattersall, 1986; Wood, 1991a, 1994; Rightmire, 1998b; Gabunia et al., 2000a; Schwartz and Tattersall, 2000), in no small part because the original diagnosis provided no comparison with the Asian fossil record}}</ref><ref>{{Cite journal | author=Suwa G; Asfaw B; Haile-Selassie Y; White T; Katoh S; WoldeGabriel G; Hart W; Nakaya H; Beyene Y| doi = 10.1537/ase.061203 | title = Early Pleistocene Homo erectus fossils from Konso, southern Ethiopia | journal = Anthropological Science | volume = 115 | issue = 2 | pages = 133 | year = 2007 | pmid = | pmc = }}</ref> |
|||
''H. erectus'' had a more modern gait and body proportions, and was the first human species to have exhibited a flat face, prominent nose, and possibly sparse body hair coverage. Though the species' brain size certainly exceeds that of ancestor species, capacity varied widely depending on the population. In earlier populations, brain development seemed to cease early in childhood, suggesting that offspring matured faster, thus limiting cognitive development. ''H. erectus'' was an [[apex predator]];<ref name="Ben-Dor2021">{{cite journal | vauthors = Ben-Dor M, Sirtoli R, Barkai R | title = The evolution of the human trophic level during the Pleistocene | journal = American Journal of Physical Anthropology | volume = 175 | issue = Suppl 72 | pages = 27–56 | date = August 2021 | pmid = 33675083 | doi = 10.1002/ajpa.24247 | doi-access = free }}</ref> sites generally show consumption of medium to large animals, such as [[bovine]]s or [[elephant]]s, and suggest the development of predatory behavior and coordinated hunting. ''H. erectus'' is associated with the [[Acheulean]] stone tool [[industry (archaeology)|industry]], and is postulated to have been the earliest human ancestor capable of using fire,<ref>{{Cite journal |last1=Zohar |first1=Irit |last2=Alperson-Afil |first2=Nira |last3=Goren-Inbar |first3=Naama |last4=Prévost |first4=Marion |last5=Tütken |first5=Thomas |last6=Sisma-Ventura |first6=Guy |last7=Hershkovitz |first7=Israel |last8=Najorka |first8=Jens |date=2022-11-14 |title=Evidence for the cooking of fish 780,000 years ago at Gesher Benot Ya'aqov, Israel |url=https://www.nature.com/articles/s41559-022-01910-z |journal=Nature Ecology & Evolution |volume=6 |issue=12 |pages=2016–2028 |language=en |doi=10.1038/s41559-022-01910-z |pmid=36376603 |bibcode=2022NatEE...6.2016Z |s2cid=253522354 |issn=2397-334X}}</ref> hunting and gathering in coordinated groups, caring for injured or sick group members, and possibly seafaring and art (though examples of art are controversial, and are otherwise rudimentary and few and far between). Intentional seafaring has also been a controversial claim.<ref name=":6">{{Cite journal |last=Botha |first=Rudolf |date=2024-04-16 |title=Did Homo erectus Have Language? The Seafaring Inference |url=https://www.cambridge.org/core/journals/cambridge-archaeological-journal/article/did-homo-erectus-have-language-the-seafaring-inference/DDCBA3076C04C50AFE23C2BF00DC461C#sec6 |journal=Cambridge Archaeological Journal |language=en |pages=1–17 |doi=10.1017/S0959774324000118 |issn=0959-7743|doi-access=free }}</ref> |
|||
A new debate appeared in 2013, with the documentation of the [[Dmanisi#Archaeological site|Dmanisi]] [[skulls]].<ref>[http://www.nature.com/news/skull-suggests-three-early-human-species-were-one-1.13972 Skull suggests three early human species were one : Nature News & Comment]</ref> Considering the large morphological variation among all Dmanisi skulls, researchers now suggest that several early human ancestors variously classified, for example, as ''[[Homo ergaster]]'', or ''[[Homo rudolfensis]]'', and perhaps even ''[[Homo habilis]]'', should instead be designated as ''Homo erectus''.<ref name=dmanisiskull5>{{cite journal |title=A Complete Skull from Dmanisi, Georgia, and the Evolutionary Biology of Early Homo |author=David Lordkipanidze, Marcia S. Ponce de Leòn, Ann Margvelashvili, Yoel Rak, G. Philip Rightmire, Abesalom Vekua, Christoph P. E. Zollikofer |journal=Science |date=18 October 2013 |volume= 342 |issue= 6156 |pages= 326–331 |doi= 10.1126/science.1238484 }}</ref><ref name=National_Geographic>{{cite news |last=Switek |first=Brian |date=17 October 2013 |title= Beautiful Skull Spurs Debate on Human History |url=http://news.nationalgeographic.com/news/2013/10/131017-skull-human-origins-dmanisi-georgia-erectus/ |newspaper= National Geographic |accessdate=22 September 2014 }}</ref> |
|||
''H. erectus'' males and females may have been roughly the same size as each other (i.e. exhibited reduced [[sexual dimorphism]]), which could indicate [[monogamy in animals|monogamy]] in line with general trends exhibited in primates. Size, nonetheless, ranged widely from {{cvt|146–185|cm|ftin|sigfig=1}} in height and {{cvt|40–68|kg}} in weight. It is unclear if ''H. erectus'' was anatomically capable of speech, though it is postulated they communicated using some [[Origin of language|proto-language]]. |
|||
==Origin== |
|||
[[File:Homo erectus.jpg|thumb|left|''Homo erectus'', [[University of Michigan Museum of Natural History]], Ann Arbor, Michigan]] |
|||
The first hypothesis of origin is that ''Homo erectus'' rose from the [[Australopithecina]] in [[East Africa]] sometime during—or perhaps even before—the [[Early Pleistocene]] geological epoch, which itself dates to 2.58 million years ago (''see'' below, "In a new finding, in 2013..."). From there it migrated, in part, by 2.0 mya, probably as a result of broad desertifying conditions developing then in eastern and northern Africa; it joined the migrations through the [[Sahara Pump Theory|"Saharan pump"]] and dispersed around much of the [[Old World]]. The fossil record shows that its development from about 1.8 mya to one mya was widely distributed: in Africa ([[Lake Turkana]] <!------> <ref>{{cite journal|authorlink=Kendrick Frazier|author=Frazier, Kendrick |url=http://www.csicop.org/si/2006-06/leakey.html|archiveurl=http://web.archive.org/web/20090110223159/http://www.csicop.org/si/2006-06/leakey.html|archivedate=2009-01-10|title= Leakey Fights Church Campaign to Downgrade Kenya Museum’s Human Fossils|journal=Skeptical Inquirer magazine |volume =30 |issue=6|date= Nov–Dec 2006}}</ref> <!-----> and [[Olduvai Gorge]]), the [[Caucasus#Prehistory|Transcaucasus]] ([[Dmanisi#Archaeological site|Dmanisi]] in [[Georgia (country)|Georgia]]), Indonesia ([[Sangiran]], [[Central Java]] and [[Trinil]], [[East Java]]), and in Vietnam, China ([[Zhoukoudian]] and [[Lantian Man|Shaanxi]]), and India.<ref>{{Cite book | author=Prins, Harald E. L.; Walrath, Dana and McBride, Bunny | title = Evolution and prehistory: the human challenge| publisher = Wadsworth Publishing| year = 2007| page = 162| url = http://books.google.com/books?id=LfYirloa_rUC&pg=PA16| isbn = 978-0-495-38190-7}}</ref> |
|||
==Taxonomy== |
|||
The second hypothesis is that ''H. erectus'' evolved in [[Eurasia]] and then migrated to Africa. They occupied the [[Dmanisi]] site from 1.85 million to 1.77 million years ago, which was about the same time or slightly before their earliest evidence in Africa.<ref name="doi10.1073/pnas.1106638108">{{Cite journal | last1 = Ferring | first1 = R. | last2 = Oms | first2 = O. | last3 = Agusti | first3 = J. | last4 = Berna | first4 = F. | last5 = Nioradze | first5 = M. | last6 = Shelia | first6 = T. | last7 = Tappen | first7 = M. | last8 = Vekua | first8 = A. | last9 = Zhvania | first9 = D. | last10 = Lordkipanidze | first10 = D. | doi = 10.1073/pnas.1106638108 | title = Earliest human occupations at Dmanisi (Georgian Caucasus) dated to 1.85-1.78 Ma | journal = Proceedings of the National Academy of Sciences | volume = 108 | issue = 26 | pages = 10432 | year = 2011 | pmid = | pmc = }}</ref><ref>[http://www.dnaindia.com/scitech/report_new-discovery-suggests-homo-erectus-originated-from-asia_1552611 New discovery suggests Homo erectus originated from Asia]. Dnaindia.com. 8 June 2011.</ref> There are several proposed explanations of the dispersal of ''H. erectus georgicus''—including whether or not Africa is the source).<ref>{{cite journal |doi=10.1016/j.quascirev.2010.04.012|date=June 2011 |last1=Augusti |first1=Jordi |last2=Lordkipanidze |first2=David |title=How "African" was the early human dispersal out of Africa?|volume=30|issue=11–12 |pages=1338–1342 |journal=Quaternary Science Reviews}}</ref> |
|||
===Naming=== |
|||
{{further|Java Man}} |
|||
[[File:Pithecanthropus erectus-PeterMaas Naturalis.jpg|thumb|left|[[Java Man]] at [[Naturalis Biodiversity Center|Naturalis]]]] |
|||
Contrary to the view [[Charles Darwin]] expressed in his 1871 book ''[[Descent of Man]]'', many late-19th century evolutionary naturalists postulated that Asia, not Africa, was the birthplace of humankind as it is midway between Europe and America, providing optimal dispersal routes throughout the world (the [[Out of Asia theory]]). Among these was German naturalist [[Ernst Haeckel]], who argued that the first human species evolved on the now-disproven hypothetical continent "[[Lemuria (continent)|Lemuria]]" in what is now Southeast Asia, from a species he termed "''[[Pithecanthropus]] alalus''" ("speechless apeman").{{sfn|Theunissen|2012|loc=p. 6}} "Lemuria" had supposedly sunk below the [[Indian Ocean]], so no fossils could be found to prove this. Nevertheless, Haeckel's model inspired Dutch scientist [[Eugène Dubois]] to journey to the [[Dutch East Indies]]. Because no directed expedition had ever discovered human fossils (the few known had all been discovered by accident), and the economy was strained by the [[Long Depression]], the Dutch government refused to fund Dubois. In 1887, he enlisted in the [[Royal Netherlands Indies Army|Dutch East India Army]] as a medical officer, and was able to secure a post in 1887 in the Indies to search for his "[[missing link (human evolution)|missing link]]" in his spare time.{{sfn|Theunissen|2012|loc=p. 33}} On [[Java]], he found a skullcap in 1891 and a [[femur]] in 1892 ([[Java Man]]) dating to the [[late Pliocene]] or [[early Pleistocene]] at the [[Trinil]] site along the [[Solo River]], which he named ''Pithecanthropus erectus'' ("upright apeman") in 1893. He attempted unsuccessfully to convince the European scientific community that he had found an upright-walking ape-man. Given few fossils of ancient humans had even been discovered at the time, they largely dismissed his findings as a malformed non-human ape.<ref name=HsiaoPei2014>{{cite journal | vauthors = Yen HP | title = Evolutionary Asiacentrism, Peking man, and the origins of sinocentric ethno-nationalism | journal = Journal of the History of Biology | volume = 47 | issue = 4 | pages = 585–625 | year = 2014 | pmid = 24771020 | doi = 10.1007/s10739-014-9381-4 | s2cid = 23308894 }}</ref> |
|||
The significance of these fossils would not be realized until the 1927 discovery of what Canadian paleoanthropologist [[Davidson Black]] called "''Sinanthropus pekinensis''" (Peking Man) at the [[Zhoukoudian]] cave near [[Beijing]], China. Black lobbied across North America and Europe for funding to continue excavating the site,{{sfn|Sigmon|1981|loc=p. 64}} which has since become the most productive ''H. erectus'' site in the world.<ref name=Yang2014>{{cite book| vauthors = Yang L |year=2014|chapter=Zhoukoudian: Geography and Culture|title=Encyclopedia of Global Archaeology|pages=7961–7965|publisher=Springer Science+Business Media|isbn=978-1-4419-0466-9|doi=10.1007/978-1-4419-0465-2_1899}}</ref> Continued interest in Java led to further ''H. erectus'' fossil discoveries at Ngandong ([[Solo Man]]) in 1931, [[Mojokerto child|Mojokerto]] (Java Man) in 1936, and [[Sangiran]] (Java Man) in 1937. The Sangiran site yielded the best preserved Java Man skull.<ref>{{cite book| vauthors = Ciochon RL, Huffman OF |year=2014|chapter=Java Man|title=Encyclopedia of Global Archaeology|pages=4182–4188| veditors = Smith C |doi=10.1007/978-1-4419-0465-2_712|isbn=978-1-4419-0426-3|s2cid=241324984 }}</ref> German paleoanthropologist [[Franz Weidenreich]] provided much of the detailed description of the Chinese specimens in several monographs. The original specimens were lost during the [[Second Sino-Japanese War]] after an attempt to smuggle them out of China for safekeeping. Only [[plaster cast|casts]] remain. |
|||
==Discovery and representative fossils== |
|||
[[File:Skull of Homo erectus, Indian Museum, Kolkata.jpg|thumb|330px|Skull of Homo erectus, [[Indian Museum]]]] |
|||
The Dutch anatomist [[Eugène Dubois]] was fascinated by [[Charles Darwin|Darwin]]'s theory of evolution especially as it applied to humankind. In 1886, he set out for [[Asia]]—which then was the region accepted as the cradle of human evolution despite Darwin's theory of African origin; see {{section link|Haeckel|Research}}—to find a human ancestor. In 1891, his team discovered a human fossil on the island of [[Java]], [[Dutch East Indies]] (now [[Indonesia]]). Excavated from the bank of the [[Solo River]] at [[Trinil]], in [[East Java]], he named the species ''Pithecanthropus erectus''—from the Greek ''πίθηκος'',<ref>''pithecos''</ref> "ape", and ''ἄνθρωπος'',<ref>''anthropos''</ref> "man"—based on a skullcap (calotte) and a femur like that of ''Homo sapiens''. |
|||
Similarities between Java Man and Peking Man led [[Ernst Mayr]] to rename both as ''Homo erectus'' in 1950. Throughout much of the 20th century, anthropologists debated the role of ''H. erectus'' in [[human evolution]]. Early in the century, due in part to the discoveries at Java and Zhoukoudian, the belief that modern humans first evolved in Asia was widely accepted. A few naturalists—[[Charles Darwin]] the most prominent among them—theorized that humans' earliest ancestors were African. Darwin had pointed out that [[chimpanzee]]s and [[gorilla]]s, [[human]]s' closest relatives, evolved and exist only in Africa.<ref>{{cite book | vauthors = Darwin CR |title=The Descent of Man and Selection in Relation to Sex|url= https://archive.org/details/descentmanandse03darwgoog |publisher=John Murray |year=1871 |isbn=978-0-8014-2085-6}}</ref> Darwin did not include [[orangutan]]s among the great [[ape]]s of the Old World, likely because he thought of orangutans as primitive humans rather than apes.<ref>{{Cite journal |last1=van Wyhe |first1=John |last2=Kjærgaard |first2=Peter C. |date=2015-06-01 |title=Going the whole orang: Darwin, Wallace and the natural history of orangutans |journal=Studies in History and Philosophy of Science Part C: Studies in History and Philosophy of Biological and Biomedical Sciences |language=en |volume=51 |pages=53–63 |doi=10.1016/j.shpsc.2015.02.006 |pmid=25861859 |s2cid=20089470 |issn=1369-8486|doi-access=free }}</ref> While Darwin considered Africa as the most probable birthplace of human ancestors, he also made the following statement about the geographic location of human origins in his book ''The Descent of Man, and Selection in Relation to Sex'': "... it is useless to speculate on this subject; for two or three anthropomorphous apes, one the Dryopithecus …, existed in Europe during the Miocene age; and since so remote a period the earth has certainly undergone many great revolutions, and there has been ample time for migration on the largest scale." (1889, pp. 155–156). |
|||
Dubois' 1891 find was the first fossil of a ''Homo''-species (or any hominin species) found as result of a directed expedition and search—and which was inspired by Darwin's radical theory that humans, like all other species, evolved from ancestral species, see [[human evolution]]. (The first found and recognized human fossil was the accidental discovery of ''[[Homo sapiens neanderthalensis]]'' in 1856, see [[List of human evolution fossils]].) The Java fossil aroused much public interest. It was dubbed by the popular press as ''[[Java Man]]''; but few scientists accepted Dubois' argument that his fossil was the [[Transitional fossil|transitional form]]—the so-called [[Transitional fossil#Missing links|"missing link"]]—between apes and humans.{{Sfn|Swisher|Curtis|Lewin|2000|p=70}} ''Java Man'' is now classified as ''Homo erectus''. |
|||
In 1949, the species was reported in [[Swartkrans]] Cave, South Africa, by South African paleoanthropologists [[Robert Broom]] and [[John Talbot Robinson]], who described it as "''Telanthropus capensis''".<ref>{{cite journal | vauthors = Curnoe D | title = A review of early Homo in southern Africa focusing on cranial, mandibular and dental remains, with the description of a new species (Homo gautengensis sp. nov.) | journal = Homo | volume = 61 | issue = 3 | pages = 151–177 | date = June 2010 | pmid = 20466364 | doi = 10.1016/j.jchb.2010.04.002 }}</ref> ''Homo'' fossils have also been reported from nearby caves, but their species designation has been a tumultuous discussion. A few North African sites have additionally yielded ''H. erectus'' remains, which at first were classified as "''Atlantanthropus mauritanicus''" in 1951.{{sfn|Sigmon|1981|loc=p. 231}} Beginning in the 1970s, propelled most notably by [[Richard Leakey]], more were being unearthed in East Africa predominantly at the [[Koobi Fora]] site, Kenya, and [[Olduvai Gorge]], Tanzania.{{sfn|Sigmon|1981|loc=p. 193}} |
|||
Most of the spectacular discoveries of ''H. erectus'' next took place at the [[Zhoukoudian|Zhoukoudian Project]], now known as the [[Peking Man]] Site, in Zhoukoudian, [[China]]. This site was first discovered by [[Johan Gunnar Andersson]] in 1921<!----------><ref name="doorKnock1">{{cite news | authorlink = | title = The First Knock at the Door| url = | format = | work = | publisher = Peking Man Site Museum | pages = | page = | date = | accessdate = | quote = In the summer of 1921, Dr. J.G. Andersson and his companions discovered this richly fossiliferous deposit through the local quarry men's guide. During examination he was surprised to notice some fragments of white quartz in tabus, a mineral normally foreign in that locality. The significance of this occurrence immediately suggested itself to him and turning to his companions, he exclaimed dramatically "Here is primitive man, now all we have to do is find him!"}}</ref> <!--------->and was first excavated in 1921, which produced two human teeth.<!----------><ref name="doorKnock2">{{cite news | authorlink = | title = The First Knock at the Door| url = | format = | work = | publisher = Peking Man Site Museum | pages = | page = | date = | accessdate = | quote = For some weeks in this summer and a longer period in 1923 Dr. Otto Zdansky carried on excavations of this cave site. He accumulated an extensive collection of fossil material, including two Homo erectus teeth that were recognized in 1926. So, the cave home of Peking Man was opened to the world.}}</ref> Canadian anatomist [[Davidson Black]]'s initial description (1921) of a lower molar as belonging to a previously unknown species (which he named ''[[Sinanthropus pekinensis]]'')<!--------><ref>from ''sino-'', a combining form of the Greek ''Σίνα'', "China", and the Latinate ''pekinensis'', "of Peking"</ref> <!---->prompted widely-publicized interest. Extensive excavations followed, which altogether uncovered 200 human fossils from more than 40 individuals including five nearly complete [[Calvaria (skull)|skullcaps]].<!-----------><ref name="historyMus5">{{cite news | authorlink = | title = Review of the History | url = | format = | work = | publisher = Peking Man Site Museum | pages = | page = | date = | accessdate = | quote = During 1927-1937, abundant human and animal fossils as well as artefact were found at Peking Man Site, it made the site to be the most productive one of the Homo erectus sites of the same age all over the world. Other localities in the vicinity were also excavated almost at the same time.}}</ref> German anatomist [[Franz Weidenreich]] provided much of the detailed description of this material in several monographs published in the journal ''Palaeontologica Sinica'' (Series D). |
|||
Archaic human fossils unearthed across Europe used to be assigned to ''H. erectus'', but have since been separated as ''[[Homo heidelbergensis|H. heidelbergensis]]'' as a result of British physical anthropologist [[Chris Stringer]]'s work.<ref name=Lumley2015>{{cite journal| vauthors = de Lumley MA |year=2015|title=L'homme de Tautavel. Un ''Homo erectus'' européen évolué. ''Homo erectus tautavelensis''|trans-title=Tautavel Man. An evolved European ''Homo erectus''. ''Homo erectus tautavelensis''|language=fr|journal=L'Anthropologie|volume=119|issue=3|pages=342–344|doi=10.1016/j.anthro.2015.06.001}}</ref> |
|||
Nearly all of the original specimens were lost during [[World War II]]; however, authentic casts were made by Weidenreich which exist at the [[American Museum of Natural History]] in [[New York]] and at the [[Institute of Vertebrate Paleontology and Paleoanthropology]] in [[Beijing]], and are considered to be reliable evidence. |
|||
===Evolution=== |
|||
Throughout much of the 20th century, anthropologists debated the role of ''H. erectus'' in [[human evolution]]. Early in the century, due in part to the discoveries at Java and Zhoukoudian, it was widely accepted that modern humans first evolved in [[Asia]]. A few naturalists—Charles Darwin most prominent among them—theorized that humans' earliest ancestors were African: Darwin pointed out that chimpanzees and gorillas, humans' closest relatives, evolved and exist only in Africa.<ref>{{cite book|last=Darwin|first = Charles R.|title=The Descent of Man and Selection in Relation to Sex|publisher=John Murray|year=1871|isbn=0-8014-2085-7}}</ref> |
|||
{{further|Early human expansions out of Africa}} |
|||
{{Human timeline}} |
|||
[[File:Carte hachereaux.jpg|thumb|upright=1.5|Map of the distribution of Middle Pleistocene ([[Acheulean]]) [[Cleaver (tool)|cleaver]] finds]] |
|||
It has been proposed that ''H. erectus'' evolved from ''[[Homo habilis|H. habilis]]'' about 2 Mya, though this has been called into question because they coexisted for at least a half a million years. Alternatively, a group of ''H. habilis'' may have been [[reproductively isolated]], and only this group developed into ''H. erectus'' ([[cladogenesis]]).<ref name=Spoor2007/> |
|||
Because the earliest remains of ''H. erectus'' are found in both Africa and East Asia (in China as early as 2.1 Mya,<ref name = "Zhu_2018">{{cite journal | vauthors = Zhu Z, Dennell R, Huang W, Wu Y, Qiu S, Yang S, Rao Z, Hou Y, Xie J, Han J, Ouyang T | display-authors = 6 | title = Hominin occupation of the Chinese Loess Plateau since about 2.1 million years ago | journal = Nature | volume = 559 | issue = 7715 | pages = 608–612 | date = July 2018 | pmid = 29995848 | doi = 10.1038/s41586-018-0299-4 | s2cid = 49670311 | author10 = Han Jiangwei ({{lang|zh-Hans|韩江伟}}) | author11 = Ouyang Tingping ({{lang|zh-Hans|欧阳婷萍}}) | author4 = Wu Yi ({{lang|zh-Hans|吴翼}}) | author5 = Qiu Shifan ({{lang|zh-Hans|邱世藩}}) | author6 = Yang Shixia ({{lang|zh-Hans|杨石霞}}) | author7 = Rao Zhiguo ({{lang|zh-Hans|饶志国}}) | author3 = Huang Weiwen ({{lang|zh-Hans|黄慰文}}) | bibcode = 2018Natur.559..608Z | author8 = Hou Yamei ({{lang|zh-Hans|侯亚梅}}) | author9 = Xie Jiubing ({{lang|zh-Hans|谢久兵}}) | name-list-style = vanc }}</ref><ref>{{Cite journal| vauthors = Barras C |year=2018|title=Tools from China are oldest hint of human lineage outside Africa|url=https://www.nature.com/articles/d41586-018-05696-8 |journal=Nature|doi=10.1038/d41586-018-05696-8|s2cid=188286436|issn=0028-0836}}</ref><ref>{{cite journal | vauthors = Hao L, Chao Rong L, Kuman K | year = 2017 | title = Longgudong, an Early Pleistocene site in Jianshi, South China, with stratigraphic association of human teeth and lithics | journal = Science China Earth | volume = 60 | issue = 3| pages = 452–462 | doi = 10.1007/s11430-016-0181-1 | bibcode = 2017ScChD..60..452L | s2cid = 132479732 }}</ref> in South Africa 2.04 Mya<ref name=Herries/><ref>{{cite web |title=Our direct human ancestor Homo erectus is older than we thought |url=https://www.eurekalert.org/pub_releases/2020-04/uoj-odh040120.php |website=EurekAlert |publisher=[[American Association for the Advancement of Science|AAAS]]}}</ref>), it is debated where ''H. erectus'' evolved. A 2011 study suggested that it was ''H. habilis'' who reached West Asia from Africa, that early ''H. erectus'' developed there, and that early ''H. erectus'' would then have dispersed from West Asia to East Asia ([[Peking Man]]), Southeast Asia ([[Java Man]]), back to Africa (''[[Homo ergaster]]''), and to Europe ([[Tautavel Man]]), eventually evolving into modern humans in Africa.<ref>{{cite journal | vauthors = Ferring R, Oms O, Agustí J, Berna F, Nioradze M, Shelia T, Tappen M, Vekua A, Zhvania D, Lordkipanidze D | display-authors = 6 | title = Earliest human occupations at Dmanisi (Georgian Caucasus) dated to 1.85-1.78 Ma | journal = Proceedings of the National Academy of Sciences of the United States of America | volume = 108 | issue = 26 | pages = 10432–10436 | date = June 2011 | pmid = 21646521 | pmc = 3127884 | doi = 10.1073/pnas.1106638108 | doi-access = free | bibcode = 2011PNAS..10810432F }}</ref><ref>{{cite journal |doi=10.1016/j.quascirev.2010.04.012|date=June 2011 | vauthors = Augusti J, Lordkipanidze D |title=How "African" was the early human dispersal out of Africa?|volume=30|issue=11–12 |pages=1338–1342 |journal=Quaternary Science Reviews|bibcode=2011QSRv...30.1338A }}</ref> Others have suggested that ''H. erectus''/''H. ergaster'' developed in Africa, where it eventually evolved into modern humans.<ref>{{cite journal| vauthors = Rightmire GP |title= Human Evolution in the Middle Pleistocene: The Role of ''Homo heidelbergensis'' |year=1998 |journal=Evolutionary Anthropology|doi=10.1002/(sici)1520-6505(1998)6:6<218::aid-evan4>3.0.co;2-6 |volume=6 |issue=6 |pages=218–227|s2cid= 26701026 }}</ref><ref name="Asfawpmid11907576">{{cite journal | vauthors = Asfaw B, Gilbert WH, Beyene Y, Hart WK, Renne PR, WoldeGabriel G, Vrba ES, White TD | display-authors = 6 | title = Remains of Homo erectus from Bouri, Middle Awash, Ethiopia | journal = Nature | volume = 416 | issue = 6878 | pages = 317–320 | date = March 2002 | pmid = 11907576 | doi = 10.1038/416317a | s2cid = 4432263 | bibcode = 2002Natur.416..317A }}</ref> |
|||
===African genesis=== |
|||
[[File:Homo Georgicus IMG 2921.JPG|thumb|Dmanisi skull 3, Fossils skull [[D2700]] and [[D2735]] jaw, two of several found in [[Dmanisi]] in the [[Republic of Georgia|Georgian]] [[Caucasus]].]] |
|||
From the 1950s forward, numerous finds in East Africa confirmed the hypothesis of an African genesis, providing fossil evidence that the earliest [[Hominini|hominins]] originated there. It is now generally accepted that ''H. erectus'' descended from either: 1) the earliest hominin genera (such as ''[[Australopithecus]]'', and possibly ''[[Ardipithecus]]''—of which is still debated whether it is hominin or hominid); or 2) the earliest ''Homo''-species (such as ''[[Homo habilis]]'' or ''[[Homo ergaster]]''). East Africa provided [[sympatric]] coexistence for ''H. erectus'' and ''H. habilis'' for several hundred-thousand years, which tends to confirm the hypothesis that they represent separate lineages from a common ancestor; that is, the ancestral relationship between them was not [[anagenetic]], but was [[Cladogenesis|cladogenetic]], which here suggests that a subgroup population of ''habilis''—or of a common ancestor of ''habilis'' and ''erectus''—became [[reproductively isolated]] from the main-group population, eventually evolving into the new species ''Homo erectus''.<ref>{{cite journal|title=Implications of new early ''Homo'' fossils from Ileret, east of Lake Turkana, Kenya|author=F. Spoor, M. G. Leakey, P. N. Gathogo, F. H. Brown, S. C. Antón, I. McDougall, C. Kiarie, F. K. Manthi & L. N. Leakey|journal=Nature|issue= 7154|pages= 688–691|date=2007-08-09|doi=10.1038/nature05986|volume=448|pmid=17687323}} "A partial maxilla assigned to H. habilis reliably demonstrates that this species survived until later than previously recognized, making an ''anagenetic'' relationship with H. erectus unlikely" (Emphasis added).</ref> |
|||
''H. erectus'' had reached [[Sangiran]], Java, by 1.5 Mya,<ref>{{Cite journal |last1=Husson |first1=Laurent |last2=Salles |first2=Tristan |last3=Lebatard |first3=Anne-Elisabeth |last4=Zerathe |first4=Swann |last5=Braucher |first5=Régis |last6=Noerwidi |first6=Sofwan |last7=Aribowo |first7=Sonny |last8=Mallard |first8=Claire |last9=Carcaillet |first9=Julien |last10=Natawidjaja |first10=Danny H. |last11=Bourlès |first11=Didier |last12=ASTER team |last13=Aumaitre |first13=Georges |last14=Bourlès |first14=Didier |last15=Keddadouche |first15=Karim |date=2022-11-08 |title=Javanese Homo erectus on the move in SE Asia circa 1.8 Ma |journal=Scientific Reports |language=en |volume=12 |issue=1 |pages=19012 |doi=10.1038/s41598-022-23206-9 |issn=2045-2322 |pmc=9643487 |pmid=36347897|bibcode=2022NatSR..1219012H }}</ref> and a second and distinct wave of ''H. erectus'' had colonized [[Zhoukoudian]], China, about 780 [[Year#Abbreviations yr and ya|kya (thousand years ago)]]. Early teeth from Sangiran are bigger and more similar to those of basal (ancestral) Western ''H. erectus'' and ''H. habilis'' than to those of the derived Zhoukoudian ''H. erectus''. However, later Sangiran teeth seem to reduce in size, which could indicate a secondary colonization event of Java by the Zhoukoudian or some closely related population.<ref>{{cite journal | vauthors = Zaim Y, Ciochon RL, Polanski JM, Grine FE, Bettis EA, Rizal Y, Franciscus RG, Larick RR, Heizler M, Eaves KL, Marsh HE | display-authors = 6 | title = New 1.5 million-year-old Homo erectus maxilla from Sangiran (Central Java, Indonesia) | journal = Journal of Human Evolution | volume = 61 | issue = 4 | pages = 363–376 | date = October 2011 | pmid = 21783226 | doi = 10.1016/j.jhevol.2011.04.009 | bibcode = 2011JHumE..61..363Z }}</ref> |
|||
In the 1950s, archaeologists [[John T. Robinson]] and [[Robert Broom]] named ''Telanthropus capensis'';<ref name="Robinson1953">{{cite journal |author=ROBINSON JT |title=The nature of Telanthropus capensis |journal=Nature |volume=171 |issue=4340 |pages=33 |date=January 1953 |pmid=13025468 |doi=10.1038/171033a0}}</ref> Robinson had discovered a jaw fragment in 1949 in [[Swartkrans]], [[South Africa]]. Later, Simonetta proposed to re-designate it to ''Homo erectus'', and Robinson agreed.<ref name="Grine2009">{{cite book | title = The First Humans: Origin and Early Evolution of the Genus Homo | author = Frederick E. Grine; John G. Fleagle; Richard E. Leakey | date = 1 Jun 2009 | publisher = Springer | page = 7 | chapter = Chapter 2: ''Homo habilis''—A Premature Discovery: Remember by One of Its Founding Fathers, 42 Years Later}}</ref> |
|||
===Subspecies=== |
|||
In 1961, [[Yves Coppens]] discovered a skull of ''Tchadanthropus uxoris'', then the earliest fossil human discovered in north Africa.<ref name=Kalb76>{{cite book | last = Kalb| first = Jon E| title =Adventures in the Bone Trade: The Race to Discover Human Ancestors in Ethiopia's Afar Depression| publisher =[[Springer Science+Business Media|Springer]] | year = 2001 | page = 76| isbn =0-387-98742-8 |url=http://books.google.com/?id=SiWispLhG1UC&printsec=frontcover&dq=Adventures+Bone+Trade#v=onepage&q&f=false |accessdate=2010-12-02}}</ref> It was reported that the fossil "had been so eroded by wind-blown sand that it mimicked the appearance of an australopith, a primitive type of ''hominid''".<ref name=Wood2002>{{Cite journal |date=11 July 2002 |author=Wood, Bernard |title=Palaeoanthropology: Hominid revelations from Chad | journal=[[Nature (journal)|Nature]] |volume= 418 |issue= 6894 |pages= 133–135 |doi= 10.1038/418133a | url=http://www.fhuce.edu.uy/antrop/cursos/abiol/links/Artics/wood%202002.pdf | archiveurl=http://web.archive.org/web/20110717090936/http://www.fhuce.edu.uy/antrop/cursos/abiol/links/Artics/wood%202002.pdf | archivedate=2011-07-17 |accessdate=2 December 2010}}</ref><!----------> Although at first considered to be a specimen of ''H. habilis'',<ref name=Cornevin>{{cite book | last = Cornevin| first = Robert| title =Histoire de l'Afrique | publisher =Payotte | year = 1967| page = 440 | isbn = 2-228-11470-7}}</ref> ''T. uxoris'' is no longer considered a valid taxon, and has been subsumed into ''H. erectus''.<ref name=Kalb76/><ref>{{cite web | title = Mikko's Phylogeny Archive | url = http://www.fmnh.helsinki.fi/users/haaramo/Metazoa/Deuterostoma/Chordata/Synapsida/Eutheria/Primates/Hominoidea/Homo_erectus.htm | archiveurl = http://web.archive.org/web/20070106024346/http://www.fmnh.helsinki.fi/users/haaramo/Metazoa/Deuterostoma/Chordata/Synapsida/Eutheria/Primates/Hominoidea/Homo_erectus.htm | archivedate = 2007-01-06 | work=Finnish Museum of Natural History, University of Helsinki}}</ref> |
|||
"[[Wushan Man]]" was proposed as ''Homo erectus wushanensis'', but is now thought to be based upon fossilized fragments of an extinct non-hominin ape.<ref name="Ciochon">{{cite journal | vauthors = Ciochon RL | title = The mystery ape of Pleistocene Asia | journal = Nature | volume = 459 | issue = 7249 | pages = 910–911 | date = June 2009 | pmid = 19536242 | doi = 10.1038/459910a | s2cid = 205047272 | doi-access = free | bibcode = 2009Natur.459..910C }}</ref> |
|||
Since the discovery of [[Java Man]] in 1893, there has been a trend in paleoanthropology of reducing the number of proposed species of ''Homo'', to the point where ''H. erectus'' includes all early ([[Lower Paleolithic]]) forms of ''Homo'' sufficiently derived from ''[[Homo habilis|H. habilis]]'' and |
|||
In 2013, a fragment of fossilized jawbone, dated to around 2.8 million years ago, was discovered in the [[Ledi-Geraru]] research area in [[Afar Region|the Afar depression]], Ethiopia.<ref name=newscientist>{{cite web|url=http://www.newscientist.com/article/dn27079-oldest-known-member-of-human-family-found-in-ethiopia.html#.VPrw7vmsXId |title=Oldest known member of human family found in Ethiopia |publisher=''[[New Scientist]]'' |date=4 March 2015 |accessdate=2015-03-07}} |
|||
distinct from early ''[[Homo heidelbergensis|H. heidelbergensis]]'' (in Africa also known as ''[[Homo rhodesiensis|H. rhodesiensis]]'').<ref name=Kaifu2005/> It is sometimes considered as a wide-ranging, polymorphous species.<ref>{{Cite journal | doi=10.1038/nature.2013.13972| title=Skull suggests three early human species were one| year=2013| vauthors = Perkins S | journal=Nature| s2cid=88314849}}</ref> |
|||
{{cite web|last=Ghosh |first=Pallab |url=http://www.bbc.co.uk/news/science-environment-31718336 |title= 'First human' discovered in Ethiopia |publisher=bbc.co.uk |date=4 March 2015 |accessdate=7 March 2015}}</ref> The fossil is considered the earliest evidence of the ''Homo'' genus known to date, and seems to be intermediate between ''Australopithecus'' and ''H. habilis''. The individual lived just after a major [[Abrupt climate change|climate shift]] in the region, when forests and waterways were rapidly replaced by arid [[savannah]], which was a domain favored by the early hominins.<ref>"Vertebrate fossils record a faunal turnover indicative of more open and probable arid habitats than those reconstructed earlier in this region, in broad agreement with hypotheses addressing the role of environmental forcing in hominin evolution at this time." {{cite journal|url=http://www.sciencemag.org/content/early/2015/03/03/science.aaa1415 |title=Late Pliocene fossiliferous sedimentary record and the environmental context of early ''Homo'' from Afar, Ethiopia |author=Erin N. DiMaggio EN, Campisano CJ, Rowan J, Dupont-Nivet G, Deino AL|journal=[[Science (journal)|Science]] |doi=10.1126/science.aaa1415 |display-authors=etal}}</ref> |
|||
Due to such a wide range of variation, it has been suggested that the ancient ''[[Homo rudolfensis|H. rudolfensis]]'' and ''[[Homo habilis|H. habilis]]'' should be considered early varieties of ''H. erectus''.<ref name=dmanisiskull5>{{cite journal | vauthors = Lordkipanidze D, Ponce de León MS, Margvelashvili A, Rak Y, Rightmire GP, Vekua A, Zollikofer CP | title = A complete skull from Dmanisi, Georgia, and the evolutionary biology of early Homo | journal = Science | volume = 342 | issue = 6156 | pages = 326–331 | date = October 2013 | pmid = 24136960 | doi = 10.1126/science.1238484 | s2cid = 20435482 | bibcode = 2013Sci...342..326L }}</ref><ref name=National_Geographic>{{cite news | vauthors = Black R |date=17 October 2013 |title= Beautiful Skull Spurs Debate on Human History |url=https://www.nationalgeographic.com/culture/article/131017-skull-human-origins-dmanisi-georgia-erectus/ |archive-url=https://web.archive.org/web/20210606185745/https://www.nationalgeographic.com/culture/article/131017-skull-human-origins-dmanisi-georgia-erectus |url-status=dead |archive-date=6 June 2021 |work=[[National Geographic (magazine)|National Geographic]] |access-date=6 June 2021}}</ref> The primitive ''H. e. georgicus'' from [[Dmanisi skulls|Dmanisi]], Georgia has the smallest brain capacity of any known Pleistocene hominin (about 600 cc), and its inclusion in the species would greatly expand the range of variation of ''H. erectus'' to perhaps include species as ''H. rudolfensis'', ''[[Homo gautengensis|H. gautengensis]]'', ''[[Homo ergaster|H. ergaster]]'', and perhaps ''H. habilis''.<ref>{{cite news |title= Skull of Homo erectus throws story of human evolution into disarray |url= https://www.theguardian.com/science/2013/oct/17/skull-homo-erectus-human-evolution | vauthors = Sample I |work= The Guardian |date= 17 October 2013 }}</ref> However, a 2015 study suggested that ''H. georgicus'' represents an earlier, more primitive species of ''Homo'' derived from an older dispersal of hominins from Africa, with ''H. ergaster/erectus'' possibly deriving from a later dispersal.<ref name="Guimares">{{cite journal |vauthors=Giumares SW, Merino CL |title=Dmanisi hominin fossils and the problem of multiple species in the early Homo genus |journal=Nexus: The Canadian Student Journal of Anthropology |volume=23 |date=September 2015 |s2cid=73528018 |url=https://pdfs.semanticscholar.org/0f43/cf9c1c394c7af01cf10160f164cdec6dd77b.pdf |archive-url=https://web.archive.org/web/20200114162011/https://pdfs.semanticscholar.org/0f43/cf9c1c394c7af01cf10160f164cdec6dd77b.pdf |archive-date=2020-01-14 }}</ref> ''H. georgicus'' is sometimes not even regarded as ''H. erectus''.<ref>{{cite journal | vauthors = Argue D, Groves CP, Lee MS, Jungers WL | title = The affinities of Homo floresiensis based on phylogenetic analyses of cranial, dental, and postcranial characters | journal = Journal of Human Evolution | volume = 107 | pages = 107–133 | date = June 2017 | pmid = 28438318 | doi = 10.1016/j.jhevol.2017.02.006 | bibcode = 2017JHumE.107..107A }}</ref><ref>{{Cite encyclopedia| vauthors = Lordkipanidze D |title=<SCP>D</SCP> manisi |chapter=Dmanisi|date=2018-10-04|encyclopedia=The International Encyclopedia of Biological Anthropology|pages=1–4| veditors = Trevathan W, Cartmill M, Dufour D, Larsen C |publisher=John Wiley & Sons, Inc.|language=en|doi=10.1002/9781118584538.ieba0139|isbn=9781118584422|s2cid=240090147}}</ref> |
|||
===''Homo erectus georgicus''=== |
|||
[[File:Dmanissi, Georgia ; Homo georgicus 1999 discovery map.png|thumb|Location of [[Dmanisi]] discovery, Georgia]] |
|||
It is debated whether the African ''H. e. ergaster'' is a separate species (and that ''H. erectus'' evolved in Asia, then migrated to Africa),<ref name="Baab">{{cite book |vauthors=Baab K |chapter=Defining Homo erectus |title=Handbook of Paleoanthropology |edition=2 |pages=2189–2219 |chapter-url=https://www.researchgate.net/publication/283477977 |date=December 2015 |doi=10.1007/978-3-642-39979-4_73 |isbn=978-3-642-39978-7 }}</ref> or is the African form (''[[sensu lato]]'') of ''H. erectus ([[sensu stricto]])''. In the latter, ''H. ergaster'' has also been suggested to represent the immediate ancestor of ''H. erectus''.<ref>{{cite book| vauthors = Tattersall I, Schwartz J |title=Extinct Humans|year=2001|isbn=978-0-8133-3482-0|place=Boulder, Colorado |publisher=Westview/Perseus |url=https://archive.org/details/extincthumans00tatt}}{{page needed|date=December 2019}}</ref> It has also been suggested that ''H. ergaster'' instead of ''H. erectus'', or some hybrid between the two, was the immediate ancestor of other archaic humans and modern humans.{{citation needed|date=April 2020}} It has been proposed that Asian ''H. erectus'' have several unique characteristics from non-Asian populations ([[autapomorphies]]), but there is no clear consensus on what these characteristics are or if they are indeed limited to only Asia. Based on supposed derived characteristics, the 120 kya Javan ''H. e. soloensis'' has been proposed to have speciated from ''H. erectus'', as ''H. soloensis'', but this has been challenged because most of the basic cranial features are maintained.<ref>{{cite journal | vauthors = Kaifu Y, Aziz F, Indriati E, Jacob T, Kurniawan I, Baba H | title = Cranial morphology of Javanese Homo erectus: new evidence for continuous evolution, specialization, and terminal extinction | journal = Journal of Human Evolution | volume = 55 | issue = 4 | pages = 551–580 | date = October 2008 | pmid = 18635247 | doi = 10.1016/j.jhevol.2008.05.002 | bibcode = 2008JHumE..55..551K }}</ref> In a wider sense, ''H. erectus'' had mostly been replaced by ''H. heidelbergensis'' by about 300 kya, with possible late survival of ''[[Homo erectus soloensis|H. erectus soloensis]]'' in Java an estimated 117-108 kya.<ref name="Rizal" /> |
|||
'''''Homo erectus georgicus''''' is the subspecies name assigned to fossil skulls and jaws found in [[Dmanisi]], [[Georgia (country)|Georgia]]. First proposed as a separate species, it is now classified within ''H. erectus''.<ref>{{cite journal |doi=10.1126/science.1072953 |pmid=12098694 |year=2002 |author=Vekua A, Lordkipanidze D, Rightmire GP, Agusti J, Ferring R, Maisuradze G, Mouskhelishvili A, Nioradze M, De Leon MP, Tappen M, Tvalchrelidze M, Zollikofer C|title=A new skull of early Homo from Dmanisi, Georgia |volume=297 |issue=5578 |pages=85–9 |journal=Science}}</ref><ref name=nature06134>{{Cite journal | author=Lordkipanidze D, Jashashvili T, Vekua A, Ponce de León MS, Zollikofer CP, Rightmire GP, Pontzer H, Ferring R, Oms O, Tappen M, Bukhsianidze M, Agusti J, Kahlke R, Kiladze G, Martinez-Navarro B, Mouskhelishvili A, Nioradze M, Rook L| title = Postcranial evidence from early Homo from Dmanisi, Georgia | doi = 10.1038/nature06134 | journal = Nature |url=http://www.mediadesk.uzh.ch/assets/downloads/Dmanisi_press_release.pdf| volume = 449 | issue = 7160 | pages = 305–310 | year = 2007 | pmid = 17882214| pmc = }}</ref><ref>{{Cite journal |
|||
| last1 = Lordkipanidze | first1 = D. |
|||
| last2 = Vekua | first2 = A. |
|||
| last3 = Ferring | first3 = R. |
|||
| last4 = Rightmire | first4 = G. P. |
|||
| last5 = Agusti | first5 = J. |
|||
| last6 = Kiladze | first6 = G. |
|||
| last7 = Mouskhelishvili | first7 = A. |
|||
| last8 = Nioradze | first8 = M. |
|||
| last9 = Ponce De León | first9 = M. S. P. |
|||
| last10 = Tappen | first10 = M. |
|||
| last11 = Zollikofer | first11 = C. P. E. |
|||
| doi = 10.1038/434717b |
|||
| title = Anthropology: The earliest toothless hominin skull |
|||
| journal = Nature |
|||
| volume = 434 |
|||
| issue = 7034 |
|||
| pages = 717–718 |
|||
| year = 2005 |
|||
| pmid = 15815618 |
|||
| pmc = |
|||
}}</ref><!---------------> The site was discovered in 1991 by Georgian scientist [[David Lordkipanidze]]. Five skulls were excavated from 1991 forward, including a "very complete" skull in 2005. Excavations at Dmanisi have yielded 73 stone tools for cutting and chopping and 34 bone fragments from unidentified [[fauna]].<ref name="doi10.1073/pnas.1106638108"/> The fossils are about 1.8 million years old. |
|||
* ''[[Bilzingsleben (Paleolithic site)|H. e. bilzingslebenensis]]'' (Vlček 1978): Originally described from a series of skulls from Bilzingsleben, with the individual of [[Samu (fossil)|Vertesszöllös]] being referred.<ref>{{Cite journal |last=Vlček |first=Emanuel |date=1978-03-01 |title=A new discovery of Homo erectus in central Europe |url=https://www.sciencedirect.com/science/article/pii/S0047248478801158 |journal=Journal of Human Evolution |language=en |volume=7 |issue=3 |pages=239–251 |doi=10.1016/S0047-2484(78)80115-8 |bibcode=1978JHumE...7..239V |issn=0047-2484}}</ref> The material historically referred to this taxon are now affiliated with [[Neanderthal]]s and the hominins at [[Sima de los huesos|Sima de los Huesos]].<ref>{{Cite journal |last1=Arsuaga |first1=Juan-Luis |last2=Martínez |first2=Ignacio |last3=Gracia |first3=Ana |last4=Carretero |first4=José-Miguel |last5=Carbonell |first5=Eudald |date=1993 |title=Three new human skulls from the Sima de los Huesos Middle Pleistocene site in Sierra de Atapuerca, Spain |url=https://www.nature.com/articles/362534a0 |journal=Nature |language=en |volume=362 |issue=6420 |pages=534–537 |doi=10.1038/362534a0 |pmid=8464493 |bibcode=1993Natur.362..534A |s2cid=4321154 |issn=1476-4687}}</ref> |
|||
After their initial assessment, some scientists were persuaded to name the Dmanisi find as a new species, ''Homo georgicus'', which they posited as a descendant of African ''[[Homo habilis]]'' and an ancestor to Asian ''Homo erectus''. This classification, however, was not supported, and the fossil was instead designated a divergent subgroup of ''Homo erectus''.<!--------------><ref>{{cite journal|url=ftp://ftp.soest.hawaii.edu/engels/Stanley/Textbook_update/Science_300/Gibbons-03b.pdf |title=A Shrunken Head for African ''Homo erectus'' |doi=10.1126/science.300.5621.893a|year=2003|last1=Gibbons|first1=A.|journal=Science|volume=300|issue=5621|pages=893a }}</ref><ref>{{Cite journal | last1 = Tattersall | first1 = I. | last2 = Schwartz | first2 = J. H. | doi = 10.1146/annurev.earth.031208.100202 | title = Evolution of the GenusHomo | journal = Annual Review of Earth and Planetary Sciences | volume = 37 | pages = 67 | year = 2009 | pmid = | pmc = }}</ref><ref>{{Cite journal |
|||
* ''[[Boskop Man|H. e. capensis]]'' (Broom 1917): A variant of "''[[Boskop Man|Homo capensis]]''",<ref name=":12"/> a taxon erected from a skull from South Africa formally classified as a type of "[[Race (human categorization)|race]]" but is now considered a representative of the [[Khoisan]].<ref>{{Cite journal |last=Singer |first=Ronald |date=1958 |title=232. The Boskop 'Race' Problem |url=https://www.jstor.org/stable/2795854 |journal=Man |volume=58 |pages=173–178 |doi=10.2307/2795854 |jstor=2795854 |issn=0025-1496}}</ref> |
|||
| last1 = Rightmire | first1 = G. P. |
|||
* ''[[Lantian Man|H. e. chenchiawoensis]]'': A name utilized in a 2007 review of Chinese archeology; the text suggests that it and ''gongwanglingensis'' are contenders in taxonomy<ref name=":03">{{Cite book |last=李学勤 |url=https://books.google.com/books?id=KtB7iumTYcsC |title=20世纪中国学术大典: 考古学, 博物馆学 |date=2007 |publisher=福建教育出版社 |isbn=978-7-5334-3641-4 |language=zh}}</ref> (despite this name not appearing in the literature). |
|||
| last2 = Lordkipanidze | first2 = D. |
|||
* ''[[Java Man|H. e. erectus]]'' (Dubois 1891):<ref>{{Cite journal |last=E |first=Dubois |date=1891 |title=Palaeontologische onderzoekingen op Java |url=https://cir.nii.ac.jp/crid/1573387448916597760 |journal=Verslag van het Mijnwezen, 3e/4e Kwartaal |pages=12–15}}</ref> The Javanese specimens of ''H. erectus'' were once classified as a distinct subspecies in the 1970s. The [[Java Man|cranium]] from [[Trinil]] is the holotype.<ref>{{Cite book |last=Tuttle |first=Russell H. |url=https://books.google.com/books?id=ejsyIZMsC9oC&pg=PA327 |title=Paleoanthropology: Morphology and Paleoecology |date=2011-05-12 |publisher=Walter de Gruyter |isbn=978-3-11-081069-1 |language=en}}</ref> |
|||
| last3 = Vekua | first3 = A. |
|||
* ''[[Homo ergaster|H. e. ergaster]]'' (Groves and Mazák 1975): Antón and Middleton (2023) suggested that ''ergaster'' should be disused based on poor diagnoses.<ref name=":22">{{Cite journal |last1=Antón |first1=Susan C. |last2=Middleton |first2=Emily R. |date=2023-06-01 |title=Making meaning from fragmentary fossils: Early Homo in the Early to early Middle Pleistocene |journal=Journal of Human Evolution |language=en |volume=179 |pages=103307 |doi=10.1016/j.jhevol.2022.103307 |issn=0047-2484|doi-access=free |pmid=37030994 |bibcode=2023JHumE.17903307A }}</ref> The name ''Homo erectus ergaster georgicus'' was created to classify the [[Dmanisi]] population as a subspecies of ''H. e. ergaster'', but [[Binomial nomenclature|quadrinomials]] are not supported by the [[ICZN]].<ref>{{Cite journal |last1=Schwartz |first1=Jeffrey H. |last2=Tattersall |first2=Ian |last3=Chi |first3=Zhang |date=2014-04-25 |title=Comment on "A Complete Skull from Dmanisi, Georgia, and the Evolutionary Biology of Early Homo " |url=https://www.science.org/doi/10.1126/science.1250056 |journal=Science |language=en |volume=344 |issue=6182 |pages=360 |doi=10.1126/science.1250056 |pmid=24763572 |bibcode=2014Sci...344..360S |s2cid=36578190 |issn=0036-8075}}</ref> |
|||
| doi = 10.1016/j.jhevol.2005.07.009 |
|||
* ''[[Dmanisi hominins|H. e. georgicus]]'' (Gabounia 1991):<ref>{{Cite journal |last1=Gabounia |first1=Léo |last2=de Lumley |first2=Marie-Antoinette |last3=Vekua |first3=Abesalom |last4=Lordkipanidze |first4=David |last5=de Lumley |first5=Henry |date=2002-09-01 |title=Découverte d'un nouvel hominidé à Dmanissi (Transcaucasie, Géorgie) |url=https://www.sciencedirect.com/science/article/pii/S1631068302000325 |journal=Comptes Rendus Palevol |language=fr |volume=1 |issue=4 |pages=243–253 |doi=10.1016/S1631-0683(02)00032-5 |bibcode=2002CRPal...1..243G |issn=1631-0683}}</ref> This hypothetical subspecific designation unites the D2600 cranium with the remainder of the Dmanisi sample, a connection that was, at the time, controversial and was only suggested if the single-species hypothesis could be proven true.<ref>{{Cite journal |last1=Rightmire |first1=G. Philip |last2=Lordkipanidze |first2=David |last3=Vekua |first3=Abesalom |date=2006-02-01 |title=Anatomical descriptions, comparative studies and evolutionary significance of the hominin skulls from Dmanisi, Republic of Georgia |url=https://www.sciencedirect.com/science/article/pii/S0047248405001624 |journal=Journal of Human Evolution |language=en |volume=50 |issue=2 |pages=115–141 |doi=10.1016/j.jhevol.2005.07.009 |pmid=16271745 |bibcode=2006JHumE..50..115R |issn=0047-2484}}</ref> |
|||
| title = Anatomical descriptions, comparative studies and evolutionary significance of the hominin skulls from Dmanisi, Republic of Georgia |
|||
* ''[[Lantian Man|H. e. gongwanglingensis]]'': A name utilized in a 2007 review of Chinese archeology; the text suggests that it and ''chenchiawoensis'' are contenders in taxonomy.<ref name=":03" /> Rukang (1992) notes that this taxon was born in a "subspecies fever".<ref>{{Cite journal |last=Rukang |first=Wu |date=1992-06-15 |title=On the classification of subspecies of Homo |url=http://www.anthropol.ac.cn/EN/ |journal=Acta Anthropologica Sinica |language=en |volume=11 |issue=2 |pages=109 |issn=1000-3193}}</ref> |
|||
| journal = Journal of Human Evolution |
|||
* ''[[Homo habilis|H. e. habilis]]'' (Leakey, Tobias, and Napier 1964): D.R. Hughes believed that the [[Olduvai Gorge|Olduvai]] specimens were not distinct enough to be assigned to ''[[Australopithecus]]'', so he created this taxon, as an early variation of ''H. erectus''.<ref>{{Cite journal |last=Tobias |first=Phillip V. |date=1991 |title=The species Homo habilis: example of a premature discovery |url=https://www.jstor.org/stable/23735461 |journal=Annales Zoologici Fennici |volume=28 |issue=3/4 |pages=371–380 |jstor=23735461 |issn=0003-455X}}</ref> |
|||
| volume = 50 |
|||
* ''[[Homo heidelbergensis|H. e. heidelbergensis]]'' (Schoetensack 1908): This taxon was used as an alternative to standard ''H. heidelbergensis'' during the middle 20th century, interpreted as a European chronospecies of the wider Middle Pleistocene hominin morph.<ref>{{Cite journal |last1=Mounier |first1=A. |last2=Caparros |first2=M. |date=2015-10-01 |title=Le statut phylogénétique d'Homo heidelbergensis – étude cladistique des homininés du Pléistocène moyen |url=https://doi.org/10.1007/s13219-015-0127-4 |journal=BMSAP |language=fr |volume=27 |issue=3 |pages=110–134 |doi=10.1007/s13219-015-0127-4 |s2cid=17449909 |issn=1777-5469}}</ref> |
|||
| issue = 2 |
|||
* ''[[Hexian Man|H. e. hexianensis]]'' (Huang 1982): Established based on the Hexian cranium.<ref>{{Cite journal |last=W |first=Huang |date=1982 |title=Preliminary study on the fossil hominid skull and fauna of Hexian, Anhui |url=https://cir.nii.ac.jp/crid/1572543025128829184 |journal=Vertebrata PalAsiatica |volume=20 |pages=plate 1}}</ref> |
|||
| pages = 115–141 |
|||
* ''[[Samu (fossil)|H. e. hungaricus]]'' (Naddeo 2023): A Hungarian paper submitted to a [[Academic conference|conference]] lists this subspecies as an alternate name for the Vertesszöllös remains.<ref>{{Cite journal |last=Naddeo |first=Michelangelo |date=2023 |title=Az ősi magyar jelképrendszer keresése |url=https://epa.oszk.hu/01400/01445/00066/pdf/EPA01445_acta_hungarica_2023_1_0460-0492.pdf |journal=Acta Historica Hungaricus |volume=38}}</ref> |
|||
| year = 2006 |
|||
* ''[[Lantian Man|H. e. lantianensis]]'' (Ju-Kang 1964): Based on hominin fossils discovered in [[Lantian County|Lantian]], originally named as a species of ''[[Sinanthropus]]'' and then reclassified as a subspecies.<ref name="Woo1964b">{{cite journal |last=Woo |first=J.-K. |year=1964 |title=The Hominid Skull of Lantian, Shenshi |url=http://www.ivpp.cas.cn/cbw/gjzdwxb/xbwzxz/200912/P020091214552793457098.pdf |journal=Vertebrata PalAsiatica |volume=10 |issue=1}}</ref> |
|||
| pmid = 16271745 |
|||
* ''[[Olduvai Hominid 9|H. e. leakeyi]]'' (Heberer 1963): A conditional name and thus unavailable for [[Taxonomy|taxonomic]] use, once used to describe [[Olduvai Hominid 9|OH 9]]. The replacement name is ''louisleakeyi''.<ref>{{Cite journal |last=Groves |first=Colin P. |date=1999-12-01 |title=Nomenclature of African Plio-Pleistocene hominins |url=http://www.sciencedirect.com/science/article/pii/S0047248499903664 |journal=Journal of Human Evolution |language=en |volume=37 |issue=6 |pages=869–872 |doi=10.1006/jhev.1999.0366 |issn=0047-2484 |pmid=10600324|bibcode=1999JHumE..37..869G }}</ref> It received limited use as a subspecies.<ref name=":12">{{Cite journal |last=Coon |first=Carleton S. |date=1966 |title=Review of The Nomenclature of the Hominidae, including a Definitive List of Hominid Taxa |url=https://www.jstor.org/stable/41449277 |journal=Human Biology |volume=38 |issue=3 |pages=344–347 |jstor=41449277 |issn=0018-7143}}</ref> |
|||
| pmc = |
|||
* ''[[Maba Man|H. e. mapaensis]]'' (Kurth 1965): A name that was proposed for the [[Maba Man|Maba cranium]], although the use of the word 'perhaps' was interpreted by the [[Paleo Core]] database to be a conditional proposal and thus not available for valid reuse under the ICZN. Groves (1989) classified it as a subspecies of ''[[Homo sapiens]]'', and Howell (1999) did not assign the species to a genus.<ref>{{Cite web |date=2023 |title=Homo erectus mapaensis Kurth, 1965 |url=https://paleocore.org/origins/nomina/detail/218/ |access-date=2023-08-07 |website=Paleo Core |language=en}}</ref> |
|||
}}</ref><ref>{{Cite journal |
|||
* ''[[Tighennif|H. e. mauritanicus]]'' (Arambourg 1954): A subspecies that received limited use as a descriptor for the cranial and mandibular material discovered at [[Tighennif|Tighenif]].<ref name=":12" /> |
|||
| doi = 10.1126/science.288.5468.1019 |
|||
* ''[[Narmada Human|H. e. narmadensis]]'' (Sonakia 1984): The name given to the [[Narmada cranium]].<ref>{{Cite journal |last1=Sonakia |first1=Arun |last2=Kennedy |first2=Kenneth A. R. |date=1985 |title=Skull Cap of an Early Man from the Narmada Valley Alluvium (Pleistocene) of Central India |url=https://www.jstor.org/stable/678879 |journal=American Anthropologist |volume=87 |issue=3 |pages=612–616 |doi=10.1525/aa.1985.87.3.02a00060 |jstor=678879 |issn=0002-7294}}</ref> |
|||
| last1 = Gabunia | first1 = L. |
|||
* ''[[Sambungmacan crania|H. e. newyorkensis]]'' (Laitman and Tattersall 2001): A name based on the Sambungmacan 3 cranium.<ref>{{Cite journal |last1=Laitman |first1=Jeffrey T. |last2=Tattersall |first2=Ian |date=2001-04-01 |title=Homo erectus newyorkensis: An Indonesian fossil rediscovered in Manhattan sheds light on the middle phase of human evolution |url=https://onlinelibrary.wiley.com/doi/10.1002/ar.1042 |journal=The Anatomical Record |language=en |volume=262 |issue=4 |pages=341–343 |doi=10.1002/ar.1042 |pmid=11275967 |s2cid=35310160 |issn=0003-276X}}</ref> |
|||
| last2 = Vekua | first2 = A. |
|||
* [[Solo Man|''H. e. ngandongensis'']] (Sartono 1976): A name that was used in the process of splitting Pithecanthropus into many subspecies.<ref>{{Cite book |last=SARTONO R |url=https://pascal-francis.inist.fr/vibad/index.php?action=getRecordDetail&idt=12954719 |title=The Javanese Pleistocene Hominids: A Re-Appraisal |date=1976}}</ref> |
|||
| last3 = Lordkipanidze | first3 = D. |
|||
* ''[[Olduvai Hominid 9|H. e. olduvaiensis]]'': A subspecies that described the OH 9 cranium, compared to the Bilzingsleben cranial fragments.<ref>{{Cite journal |last1=Vlček |first1=Emanuel |last2=Mania |first2=Dietrich |date=1977 |title=Ein Neuer Fund Von Homo Erectus in Europa: Bilzingsleben (ddr) |url=https://www.jstor.org/stable/26294532 |journal=Anthropologie (1962-) |volume=15 |issue=2/3 |pages=159–169 |jstor=26294532 |issn=0323-1119}}</ref> |
|||
| last4 = Swisher Cc | first4 = 3. |
|||
* ''[[Peking Man|H. e. pekinensis]]'' (Black and Zdansky 1927): Originally assigned the type of ''Sinanthropus'' based on a single molar.<ref>{{Cite journal |last=D |first=Black |date=1927 |title=On a lower molar hominid tooth from the Chou Kou Tien deposit |url=https://cir.nii.ac.jp/crid/1571698599180803456 |journal=Palaeont. Sinica, Ser. D |volume=7 |pages=1–29}}</ref> Antón and Middleton (2023) suggested that [[Zhoukoudian Peking Man Site|Zhoukoudian]] and Nanjing may be referrable under this name if they exhibit enough discontinuity from ''H. erectus'' proper.<ref name=":22" /> |
|||
| last5 = Ferring | first5 = R. |
|||
* ''[[Reilingen cranium|H. e. reilingensis]]'' (Czarnetzki 1989): Referring to a single cranial fragment, this subspecies is now considered a member of the Neanderthal lineage.<ref>{{Cite journal |last1=Dean |first1=David |last2=Hublin |first2=Jean-Jacques |last3=Holloway |first3=Ralph |last4=Ziegler |first4=Reinhard |date=1998 |title=On the phylogenetic position of the pre-Neandertal specimen from Reilingen, Germany |journal=Journal of Human Evolution |language=en |volume=34 |issue=5 |pages=485–508 |doi=10.1006/jhev.1998.0214|doi-access=free |pmid=9614635 |bibcode=1998JHumE..34..485D }}</ref> |
|||
| last6 = Justus | first6 = A. |
|||
* ''[[Solo Man|H. e. soloensis]]'' (Oppenoorth 1932): The original name devised by Oppenoorth for the Ngandong crania.<ref>{{Cite journal |last=Oppenoorth |first=W.F.F. |date=1932 |title=Homo (Javanthropus) soloensis. Ein plistocene mensch van Java |journal=Wetenschappelijke Mededeelingen |volume=20 |pages=49–75}}</ref> |
|||
| last7 = Nioradze | first7 = M. |
|||
* ''[[Tautavel Man|H. e. tautavelensis]]'' (de Lumley and de Lumley 1971): Referring to the remains discovered at [[Caune de l'Arago|Arago]], with many preferring allocation to ''Homo heidelbergensis''.<ref>{{Cite thesis |title=Reconstitution et position phylétique des restes crâniens de l'Homme de Tautavel (Arago 21-47) et de Biache-Saint-Vaast 2. Apports de l'imagerie et de l'analyse tridimensionnelle. |url=https://theses.hal.science/tel-00653112 |publisher=Université Paul Cézanne - Aix-Marseille III |date=2005-11-30 |degree=phdthesis |language=fr |first=Gaspard |last=Guipert}}</ref> The remains were determined not to be ''H. erectus'' by Antón and Middleton (2023).<ref name=":22" /> |
|||
| last8 = Tvalchrelidze | first8 = M. |
|||
* ''[[Java Man|H. e. trinilensis]]'' (Sartono 1976): A tentative classification scheme, thus making the name conditional and unable for use.<ref>{{Cite journal |last=Sartono |first=S. |date=1980 |title=On the Javanese Pleistocene hominids: A reappraisal |journal=Abstracts of the IUSPP Nice}}</ref> |
|||
| last9 = Antón | first9 = S. C. |
|||
* ''[[Wajak crania|H. e. wadjakensis]]'' (Dubois 1921): A species established by Eugene Dubois based on the Wajak skulls.<ref>{{Cite journal |last=Dubois |first=E. |date=1921-01-01 |title=The proto-Australian fossil man of Wadjak, Java |url=https://ui.adsabs.harvard.edu/abs/1921KNAB...23.1013D |journal=Koninklijke Nederlandse Akademie van Wetenschappen Proceedings Series B Physical Sciences |volume=23 |pages=1013–1051|bibcode=1921KNAB...23.1013D }}</ref> Pramujiono classified these materials as a subspecies, and incorrectly self-published the name as ''wajakensis''.<ref>{{Cite journal |last=Pramujiono |first=Agung |title=BERBAGAI PANDANGAN ASAL BANGSA DAN BAHASA INDONESIA: DARI KAJIAN LINGUSITIK HISTORIS KOMPARATIF SAMPAI ARKEOLINGUISTIK DAN PALEOLINGUISTIK |url=https://www.academia.edu/9091394 |website=Academia.edu}}</ref> |
|||
| last10 = Bosinski | first10 = G. |
|||
* ''[[Wushan Man|H. e. wushanensis]]'' (Huang and Fang 1991): Originally conceived as a hominin, the remains this taxon is founded on are more likely referred to [[Ponginae]].<ref>{{Cite journal |last1=Wei |first1=Guangbiao |last2=Huang |first2=Wanbo |last3=Chen |first3=Shaokun |last4=He |first4=Cunding |last5=Pang |first5=Libo |last6=Wu |first6=Yan |date=2014-12-15 |title=Paleolithic culture of Longgupo and its creators |url=https://www.sciencedirect.com/science/article/pii/S1040618214002110 |journal=Quaternary International |series=Multidisciplinary Perspectives on the Gigantopithecus Fauna and Quaternary Biostratigraphy in East Asia |language=en |volume=354 |pages=154–161 |doi=10.1016/j.quaint.2014.04.003 |bibcode=2014QuInt.354..154W |issn=1040-6182}}</ref><ref>{{Cite journal |last1=Zanolli |first1=Clément |last2=Kullmer |first2=Ottmar |last3=Kelley |first3=Jay |last4=Bacon |first4=Anne-Marie |last5=Demeter |first5=Fabrice |last6=Dumoncel |first6=Jean |last7=Fiorenza |first7=Luca |last8=Grine |first8=Frederick E. |last9=Hublin |first9=Jean-Jacques |last10=Nguyen |first10=Anh Tuan |last11=Nguyen |first11=Thi Mai Huong |last12=Pan |first12=Lei |last13=Schillinger |first13=Burkhard |last14=Schrenk |first14=Friedemann |last15=Skinner |first15=Matthew M. |date=May 2019 |title=Evidence for increased hominid diversity in the Early to Middle Pleistocene of Indonesia |url=https://www.nature.com/articles/s41559-019-0860-z |journal=Nature Ecology & Evolution |language=en |volume=3 |issue=5 |pages=755–764 |doi=10.1038/s41559-019-0860-z |pmid=30962558 |bibcode=2019NatEE...3..755Z |s2cid=102353734 |issn=2397-334X}}</ref> |
|||
| last11 = Jöris | first11 = O. |
|||
* ''[[Yuanmou Man|H. e. yuanmouensis]]'' (Li ''et al.'' 1977): Based on hominin remains<ref>{{Cite journal |last=Chengzhi |first=Hu |date=1973 |title=Yunnan Yuanmou faxian de yuanren yachi huashi |journal=Dizhi Xuebao |volume=1 |pages=65–71}}</ref> that Antón and Middleton (2023) suggest do not belong to the taxon H. erectus, although they do not provide an alternate classification.<ref name=":22" /> |
|||
| last12 = Lumley | first12 = M. A. |
|||
| last13 = Majsuradze | first13 = G. |
|||
| last14 = Mouskhelishvili | first14 = A. |
|||
| title = Earliest Pleistocene hominid cranial remains from Dmanisi, Republic of Georgia: Taxonomy, geological setting, and age |
|||
| journal = Science |
|||
| volume = 288 |
|||
| issue = 5468 |
|||
| pages = 1019–1025 |
|||
| year = 2000 |
|||
| pmid = 10807567 |
|||
}}</ref> |
|||
[[File:Homo Georgicus IMG 2921.JPG|thumb|upright|Modern reproduction of the [[Dmanisi skull 3]] (fossils skull D2700 and jaw D2735, two of several found in [[Dmanisi]] in the [[Georgia (country)|Georgian]] [[Transcaucasus]])]] |
|||
The fossil skeletons present a species primitive in its skull and upper body but with relatively advanced spine and lower limbs, inferring greater mobility than the previous morphology.<ref name="Bower, Bruce 275–276">{{cite journal|doi=10.2307/4019325|title=Evolutionary back story: Thoroughly modern spine supported human ancestor|author=Bower, Bruce|journal=Science News|volume =169|issue =18| pages =275–276|date= 3 May 2006 }}</ref> <!----->It is now thought ''not'' to be a separate species, but to represent a stage soon after the transition between ''H. habilis'' to ''H. erectus''; it has been dated at 1.8 mya.<ref name=nature06134/><ref>{{cite news |first=John Noble |last=Wilford |authorlink=John Noble Wilford |date=19 September 2007 |title=New Fossils Offer Glimpse of Human Ancestors |url=http://www.nytimes.com/2007/09/19/science/19cnd-fossil.html |work=[[The New York Times]] |accessdate=9 September 2009}}</ref> The assemblage includes one of the largest [[Pleistocene]] ''Homo'' mandibles (D2600), one of the smallest [[Lower Pleistocene]] mandibles (D211), a nearly complete sub-adult (D2735), and a toothless specimen [[Dmanisi skull 4|D3444/D3900]].<ref>{{cite journal |doi=10.1016/j.jhevol.2008.02.003 |pmid=18394678 |year=2008 |last1=Rightmire |first1=G. Philip |last2=Van Arsdale |first2=Adam P. |last3=Lordkipanidze |first3=David |authorlink3=David Lordkipanidze |title=Variation in the mandibles from Dmanisi, Georgia |volume=54 |issue=6 |pages=904–8 |journal=Journal of Human Evolution}}</ref> |
|||
===Descendants and synonyms=== |
|||
Two of the skulls—[[skull D2700|D2700]], with a brain volume of {{Convert|600|cc}}, and [[Dmanisi skull|D4500 or Dmanisi Skull 5]], with a brain volume of about 546 centimetres—present the two smallest and most primitive [[Hominina]] skulls from the Pleistocene period.<ref name=dmanisiskull5>{{cite journal |title=A Complete Skull from Dmanisi, Georgia, and the Evolutionary Biology of Early Homo |author=David Lordkipanidze1, Marcia S. Ponce de Lean, Ann Margvelashvili, Yoel Rak, G. Philip Rightmire, Abesalom Vekua, Christoph P. E. Zollikofer |journal=Science |date=18 October 2013 |volume= 342 |issue= 6156 |pages= 326–331 |doi= 10.1126/science.1238484 }}</ref> <!------------>The variation in these skulls were compared to variations in modern humans and within a sample group of chimpanzees. The researchers found that, despite appearances, the variations in the Dmanisi skulls were no greater than those seen among modern people and among chimpanzees. These findings suggest that previous fossil finds that were classified as different species on the basis of the large morphological variation among them—including ''[[Homo rudolfensis]]'', ''[[Homo gautengensis]]'', ''[[H. ergaster]]'', and potentially even ''[[H. habilis]]''—should perhaps be re-classified to the same lineage as ''Homo erectus''.<ref>{{cite news |title= Skull of Homo erectus throws story of human evolution into disarray |url= http://www.theguardian.com/science/2013/oct/17/skull-homo-erectus-human-evolution |author=Ian Sample |work= The Guardian |date= 17 October 2013 }}</ref> |
|||
{{More citations needed section|date=July 2021}} |
|||
''Homo erectus'' is the most long-lived species of ''Homo'', having survived for almost two million years. By contrast, ''[[Homo sapiens]]'' emerged about a third of a million years ago. |
|||
Regarding many [[archaic humans]], there is no definite consensus as to whether they should be classified as [[Human subspecies|subspecies]] of ''H. erectus'' or ''H. sapiens'' or as separate species. |
|||
==Classification and distinctions== |
|||
* African ''H. erectus'' candidates |
|||
[[Paleoanthropologist]]s continue to debate the classification of ''Homo erectus'' and ''[[Homo ergaster]]'' as separate species. One school of thought suggests dropping the taxon ''Homo erectus'' and equating ''H. erectus'' with the archaic ''H. sapiens''.<ref>{{cite journal|author=Weidenreich, F. |year=1943|title=The "Neanderthal Man" and the ancestors of "Homo Sapiens"|jstor=662864|doi=10.1525/aa.1943.45.1.02a00040 |journal=American Anthropologist |volume=45|pages=39–48}}</ref><ref>{{cite journal|author=Jelinek, J. |year=1978|title= Homo erectus or Homo sapiens? |journal=Rec. Adv. Primatol.|volume=3|pages=419–429}}</ref><ref>{{cite journal|author=Wolpoff, M.H. |year=1984|title= Evolution of Homo erectus: The question of stasis|jstor=2400612|journal= Palaeobiology |volume=10|issue=4|pages=389–406}}</ref><ref>{{cite journal|jstor=681178|author=Frayer, D.W., Wolpoff, M.H.; Thorne, A.G.; Smith, F.H. and Pope, G.G. |year=1993|title= Theories of modern human origins: The paleontological test|journal=American Anthropologist|volume=95|pages=14–50|doi=10.1525/aa.1993.95.1.02a00020}}</ref><!---------> Another calls ''H. ergaster'' the direct African ancestor of ''H. erectus'', proposing that ''erectus'' emigrated out of Africa to [[Asia]] while branching into a distinct species.<ref>{{cite book|last=Tattersall|first=Ian and Jeffrey Schwartz|title=Extinct Humans|year=2001|isbn=0-8133-3482-9|place=Boulder, Colorado|publisher= Westview/Perseus}}</ref><!---------> Some scholars dispense with the species name ''ergaster'', making no distinction between such fossils as the [[Turkana Boy]] and [[Peking Man]].{{Citation needed|date=January 2010}} Still, "''Homo ergaster''" has gained some acceptance as a valid taxon, and the two species are still usually defined as distinct African and Asian [[populations]] of the greater species ''H. erectus'', that is, "''Homo erectus [[sensu lato]]''". |
|||
** ''[[Homo ergaster]]'' (or "African ''H. erectus''") |
|||
** ''[[Homo naledi]]'' |
|||
* Eurasian ''H. erectus'' candidates: |
|||
** ''[[Homo antecessor]]'' |
|||
** ''[[Homo heidelbergensis]]'' |
|||
* ''[[Homo floresiensis]]''<ref>There was long-standing uncertainty whether ''H. floresiensis'' should be considered close to ''H. erectus'', close to ''H. sapiens'', or an altogether separate species. |
|||
In 2017, it was suggested on morphological grounds that ''H. floresiensis'' is a sister species to either ''H. habilis'' or to a minimally ''habilis''-''erectus''-''ergaster''-''sapiens'' [[clade]], and its line much more ancient than Homo erectus itself. |
|||
{{cite journal | vauthors = Argue D, Groves CP, Lee MS, Jungers WL | title = The affinities of Homo floresiensis based on phylogenetic analyses of cranial, dental, and postcranial characters | journal = Journal of Human Evolution | volume = 107 | pages = 107–133 | date = June 2017 | pmid = 28438318 | doi = 10.1016/j.jhevol.2017.02.006 | bibcode = 2017JHumE.107..107A }}</ref> |
|||
* ''[[Homo rhodesiensis]]'' |
|||
* the Narmada fossil, discovered in 1982 in [[Madhya Pradesh]], India, was at first suggested as ''H. erectus'' or ''Homo erectus narmadensis''.<ref>{{cite journal | vauthors = Kennedy KA, Sonakia A, Chiment J, Verma KK | title = Is the Narmada hominid an Indian Homo erectus? | journal = American Journal of Physical Anthropology | volume = 86 | issue = 4 | pages = 475–496 | date = December 1991 | pmid = 1776655 | doi = 10.1002/ajpa.1330860404 }}</ref> |
|||
''[[Meganthropus]]'', based on fossils found in Java, dated to between 1.4 and 0.9 Mya, was tentatively grouped with ''H. erectus'' in contrast to earlier interpretations of it as a giant species of early human<ref name=Kaifu2005>{{cite journal | vauthors = Kaifu Y, Baba H, Aziz F, Indriati E, Schrenk F, Jacob T | title = Taxonomic affinities and evolutionary history of the Early Pleistocene hominids of Java: dentognathic evidence | journal = American Journal of Physical Anthropology | volume = 128 | issue = 4 | pages = 709–726 | date = December 2005 | pmid = 15761880 | doi = 10.1002/ajpa.10425 }}</ref> although older literature has placed the fossils outside of ''Homo'' altogether.<ref>Krantz, G.S. (1975). "An explanation for the diastema of Javan erectus Skull IV". In: Paleoanthropology, Morphology and Paleoecology. La Hague: Mouton, 361–372.</ref> However, Zanolli et al. (2019) judged ''Meganthropus'' to be a distinct genus of extinct ape.<ref>{{cite journal | vauthors = Zanolli C, Kullmer O, Kelley J, Bacon AM, Demeter F, Dumoncel J, Fiorenza L, Grine FE, Hublin JJ, Nguyen AT, Nguyen TM, Pan L, Schillinger B, Schrenk F, Skinner MM, Ji X, Macchiarelli R | display-authors = 6 | title = Evidence for increased hominid diversity in the Early to Middle Pleistocene of Indonesia | journal = Nature Ecology & Evolution | volume = 3 | issue = 5 | pages = 755–764 | date = May 2019 | pmid = 30962558 | doi = 10.1038/s41559-019-0860-z | bibcode = 2019NatEE...3..755Z | s2cid = 102353734 | url = https://kar.kent.ac.uk/72814/1/01-Indonesian_hominid_paleobiodiversity_v2.pdf }}</ref> |
|||
Some have insisted that [[Ernst Mayr]]'s [[Biological species concept|biological species definition]] cannot be used to test the above hypotheses—that is, that the two species might be considered the same. Alternatively, the amount of variation of cranial morphology between known specimens of ''H. erectus'' and ''H. ergaster'' can be compared to the same variation within an appropriate population of living primates (that is, one of similar geographical distribution or close evolutionary relationship), such that: if the amount of variation between ''H. erectus'' and ''H. ergaster'' is greater than that within an appropriately selected population, for example, say, [[macaque]]s, then ''H. erectus'' and ''H. ergaster'' may be considered as two different species. |
|||
==Anatomy== |
|||
Finding an extant (i.e., living) model suitable for field study, analysis, and comparison is very important; and selecting a living sample population of an appropriate species can be difficult. (For example, the morphological variation among the global population of ''H. sapiens'' is small,<ref name="Java Man"/> so our own species diversity may not be a trustworthy comparison. Fossils found in [[Dmanisi]], [[Georgia (country)|Georgia]] were originally designated as a separate (but closely related) species; but subsequent specimens showed their variation to be within the range of ''Homo erectus''. and they are now classified as ''Homo erectus georgicus''.) |
|||
===Head=== |
|||
[[File:Skull pekingman.jpg|thumb|left|upright=0.8|Skull of ''H. e. pekinensis'' showing a flat face, pronounced brow ridge, and a [[sagittal keel]]]] |
|||
''Homo erectus'' featured a flat face compared to earlier hominins; pronounced brow ridge; and a low, flat skull.<ref>{{cite journal | vauthors = Baba H, Aziz F, Kaifu Y, Suwa G, Kono RT, Jacob T | title = Homo erectus calvarium from the Pleistocene of Java | journal = Science | volume = 299 | issue = 5611 | pages = 1384–1388 | date = February 2003 | pmid = 12610302 | doi = 10.1126/science.1081676 | s2cid = 20437090 }}</ref><ref name=Balzeau2006/> The presence of [[sagittal keel|sagittal]], [[frontal suture|frontal]], and [[coronal suture|coronal]] keels, which are small crests that run along these [[suture (anatomy)|suture]] lines, has been proposed to be evidence of significant thickening of the skull, specifically the [[cranial vault]]. [[CT scan]] analyses reveal this to not be the case. However, the [[squamous part of occipital bone]], particularly the [[internal occipital crest]], at the rear of the skull is notably thicker than that of modern humans, likely a [[basal (phylogenetics)|basal]] (ancestral) trait.<ref name=Balzeau2006>{{cite journal| vauthors = Balzeau A |year=2006|title=Are thickened cranial bones and equal participation of the three structural bone layers autapomorphic traits of ''Homo erectus''?|journal=Bulletins et mémoires de la Société d'Anthropologie de Paris|volume=18|issue=3–4|pages=145–163|doi=10.4000/bmsap.1528|url=https://journals.openedition.org/bmsap/1528}}</ref><ref>{{cite journal | vauthors = Copes LE, Kimbel WH | title = Cranial vault thickness in primates: Homo erectus does not have uniquely thick vault bones | journal = Journal of Human Evolution | volume = 90 | pages = 120–134 | date = January 2016 | pmid = 26767964 | doi = 10.1016/j.jhevol.2015.08.008 | doi-access = free | bibcode = 2016JHumE..90..120C }}</ref> The fossil record indicates that ''H. erectus'' was the first human species to have featured a projecting nose, which is generally thought to have evolved in response to breathing dry air in order to retain moisture.<ref>{{cite journal | vauthors = Franciscus RG, Trinkaus E | title = Nasal morphology and the emergence of Homo erectus | journal = American Journal of Physical Anthropology | volume = 75 | issue = 4 | pages = 517–527 | date = April 1988 | pmid = 3133950 | doi = 10.1002/ajpa.1330750409 | author2-link = Erik Trinkaus }}</ref> American psychologist Lucia Jacobs hypothesized that the projecting nose instead allowed for distinguishing the direction different smells come from (stereo olfaction) to facilitate navigation and long-distance migration.<ref>{{cite journal | vauthors = Jacobs LF | title = The navigational nose: a new hypothesis for the function of the human external pyramid | journal = The Journal of Experimental Biology | volume = 222 | issue = Pt Suppl 1 | page = jeb186924 | date = February 2019 | pmid = 30728230 | doi = 10.1242/jeb.186924 | doi-access = free }}</ref> |
|||
The average brain size of Asian ''H. erectus'' is about {{cvt|1000|cc}}. However, markedly smaller specimens have been found in Dmanisi, Georgia (''H. e. georgicus''); [[Koobi Fora]] and [[Olorgesailie]], Kenya; and possibly [[Gona, Ethiopia|Gona]], Ethiopia. Overall, ''H. erectus'' brain size varies from {{cvt|546–1251|cc}},<ref name=Anton2016/> which is greater than the range of variation seen in modern humans and chimps, though less than that of gorillas.{{Citation needed|date=August 2020}} |
|||
''H. erectus'' fossils show a [[Human cranium|cranial]] capacity greater than that of ''[[Homo habilis]]'' (although the Dmanisi specimens have distinctively small crania): the earliest fossils show a cranial capacity of 850 cm³, while later Javan specimens measure up to 1100 cm³,<ref name="Java Man">Swisher, Carl Celso III; Curtis, Garniss H. and Lewin, Roger (2002) ''Java Man'', Abacus, ISBN 0-349-11473-0.</ref> overlapping that of ''H. sapiens''.; the [[frontal bone]] is less sloped and the dental arcade smaller than that of the [[australopithecine]]s; the face is more orthognatic (less protrusive) than either the australopithecines or ''H. habilis'', with large brow-ridges and less prominent [[zygoma]]ta (cheekbones). The early hominins stood about {{height|m=1.79|precision=0}}<ref name = Bryson>{{cite book |author=Bryson, Bill |title=A Short History of Nearly Everything: Special Illustrated Edition |publisher=Doubleday Canada |location=Toronto |year= 2005|isbn=0-385-66198-3}}</ref>—only 17 percent of modern male humans are taller<!------><ref name = Khanna>{{cite book |author=Khanna, Dev Raj |title=Human Evolution |publisher=Discovery Publishing House |year= 2004|page=195 |isbn=978-8171417759|url=http://books.google.com/books?id=aTxkAcdYgu0C&pg=PA195 |quote=African H. erectus, with a mean stature of 170 cm, would be in the tallest 17 percent of modern populations, even if we make comparisons only with males |accessdate=30 March 2013 }}</ref>—and were extraordinarily slender, with long arms and legs.<!-----><ref name = Roylance>{{cite news |title=A Kid Tall For His Age |author=Roylance, Frank D. Roylance |url=http://articles.baltimoresun.com/1994-02-06/news/1994037060_1_erectus-skeleton-neanderthal |newspaper=Baltimore Sun |quote=Clearly this population of early people were tall, and fit. Their long bones were very strong. We believe their activity level was much higher than we can imagine today. We can hardly find Olympic athletes with the stature of these people |date=6 February 1994 |accessdate=30 March 2013}}</ref> |
|||
[[File:Homo erectus reconstruction, Natural History Museum, London.jpg|thumb|upright=1.5|Homo erectus reconstruction, Natural History Museum, London.]] |
|||
[[Sexual dimorphism]] in ''H. erectus''—males are about 25% larger than females—is slightly greater than seen in the later ''H. sapiens'', but less than that of the earlier genus ''[[Australopithecus]]''. Regarding evolution of human physiology, the discovery of the skeleton of "[[Turkana boy]]" (''Homo ergaster'') near [[Lake Turkana]], [[Kenya]], by [[Richard Leakey]] and [[Kamoya Kimeu]] in 1984—one of the most complete hominin skeletons ever discovered—has contributed greatly to the interpretation. |
|||
In an article published in 2021 titled "Interpopulational variation in human brain size: Implications for hominin cognitive phylogeny," it was found that the brain size of Asian ''H. erectus'' over the last 600,000 years overlaps significantly with modern human populations. Significantly, some small brained modern populations showed greater affinity with ''H. erectus'' than they did with other large brained and large bodied modern populations. The paper points out methodological flaws in current understanding of brain size increase in human evolution, where species averages are compared with fossils, which overlooks interpopulational variation. It also overlooks the fact that some modern populations have not seen any dramatic brain size increase relative to ''H. erectus'' with most of the increase occurring in northern populations, which has the result of obscuring interpopulational variation. As the authors write '...the increase in the mean of ''H. sapiens'' cranial capacity is to a large extent due to an increase in the upper limit with a much less pronounced increase in the lower limit relative to our ''H. erectus'' sample. And this increase in the upper limit seems to be more pronounced in northern populations – which may be a result of correlated increases in body size in addition to climatic factors. Consequently, the authors argue that purely based on brain size similarities, Asian ''H. erectus'' could be re-classified as a subspecies of ''H. sapiens'', that is ''H. sapiens soloensis'' - as was suggested by earlier authors.<ref>{{Cite journal|doi = 10.2478/anre-2021-0029|title = Interpopulational variation in human brain size: Implications for hominin cognitive phylogeny|year = 2022|last1 = Clark|first1 = Gary|last2 = Henneberg|first2 = Maciej|journal = Anthropological Review|volume = 84|issue = 4|pages = 405–429|doi-access = free}}</ref> |
|||
Dentally, ''H. erectus'' have the thinnest [[tooth enamel|enamel]] of any Plio–Pleistocene hominin. Enamel prevents the tooth from breaking from hard foods but impedes shearing through tough foods. The [[Mandible#Body|bodies of the mandibles]] of ''H. erectus'', and all early ''Homo'', are thicker than those of modern humans and all living apes. The mandibular body resists torsion from the [[bite force]] or chewing, meaning their jaws could produce unusually powerful stresses while eating, but the practical application of this is unclear. Nonetheless, the mandibular bodies of ''H. erectus'' are somewhat thinner than those of early ''Homo''. The premolars and molars also have a higher frequency of pits than ''H. habilis'', suggesting ''H. erectus'' ate more brittle foods (which cause pitting). These all indicate that the ''H. erectus'' mouth was less capable of processing hard foods and more at shearing through tougher foods, thus reducing the variety of foods it could process, likely as a response to tool use.<ref name=Ungar2006>{{cite journal| vauthors = Ungar PS, Grine FE |year=2006|title=Diet in Early ''Homo'': A Review of the Evidence and a New Model of Adaptive Versatility|journal=Annual Review of Anthropology|volume=35|pages=208–228|doi=10.1146/annurev.anthro.35.081705.123153}}</ref> |
|||
===Interpreting evolution: ''H. erectus'' / ''H. ergaster'' / ''H. sapiens''=== |
|||
[[File:Homo-Stammbaum, Version Stringer-en.svg|425px|thumb|Springer graph-model of the evolution of several species of genus ''Homo'' over the last 2 million years (vertical axis). The rapid "[[Recent African origin of modern humans|Out of Africa]]" expansion of ''H. sapiens'' is indicated at the top of the diagram.<ref name="Stringer 2012 33–35">{{cite journal | last=Stringer | first=C. | title=What makes a modern human | journal=Nature | year=2012 | volume=485 | issue=7396 | pages=33–35 | doi=10.1038/485033a | pmid=22552077}}</ref>]] |
|||
[[File:Human evolution chart-en.svg|An alternate graph-model of the temporal and geographical distribution of several ''Homo'' species, evolving over the last two million years ; proposed by Reed, et al., redrawn from Springer.<ref name="ncbi.nlm.nih.gov">[https://www.ncbi.nlm.nih.gov/pmc/articles/PMC521174/figure/pbio-0020340-g005/ "Figure 5. Temporal and Geographical Distribution of Hominid Populations Redrawn from Stringer (2003)"] (edited from source), in {{cite journal |last1=Reed |first1=David L. |last2=Smith |first2=Vincent S. |last3=Hammond |first3=Shaless L. |last4=Rogers |first4=Alan R. |last5=Clayton |first5=Dale H. |date=November 2004 |title=Genetic Analysis of Lice Supports Direct Contact between Modern and Archaic Humans |journal=[[PLOS Biology]] |location=San Francisco, CA |publisher=[[PLOS]] |volume=2 |issue=11 |page=e340 |doi=10.1371/journal.pbio.0020340 |issn=1545-7885 |pmc=521174 |pmid=15502871 |display-authors=3}}</ref> Note the depiction of ''Homo ergaster'' as an ancestor of ''Homo erectus''.|thumb|425px]] |
|||
===Body=== |
|||
Springer (2003, 2012) and Reed, et al. (2004) and others have produced schematic graph-models for interpreting the evolution of ''Homo sapiens'' from earlier species of ''Homo'', including ''Homo erectus'' and/or ''Homo ergaster'', ''see'' graphs at right. Blue areas denote the existence of one or more hominin species at a given time and place (that is, region). These and other interpretations differ mainly in the taxonomy and geographical distribution of species.<ref name="Stringer 2012 33–35"/><!-----><ref name="ncbi.nlm.nih.gov"/> |
|||
[[File:Turkana boy by Mauricio Antón.jpg|thumb|Skeleton and reconstruction of [[Turkana Boy]] by [[Mauricio Antón]]]] |
|||
Like modern humans, ''H. erectus'' varied widely in size, ranging from {{cvt|146–185|cm|ftin|sigfig=1}} in height and {{cvt|40–68|kg}} in weight, thought to be due to regional differences in climate, mortality rates, or nutrition.<ref>{{cite book |
|||
| isbn = 9780674600751 |
|||
| url = https://books.google.com/books?id=RGGVsCPuv1cC&dq=homo+erectus+tall&pg=PA412 |
|||
| title = The Nariokotome Homo erectus skeleton |
|||
| publisher = Harvard University Press |
|||
| date = 1993 |
|||
| access-date = 2 October 2022 |
|||
| page = 412 |
|||
| author = Alan Walker, Richard Leakey |
|||
}}</ref><ref>{{cite journal| vauthors = Migliano AB, Guillon M |year=2012|title=The Effects of Mortality, Subsistence, and Ecology on Human Adult Height and Implications for ''Homo'' Evolution|journal=Current Anthropology|volume=53|issue=S6|pages=359–368|url=https://www.researchgate.net/publication/233905432|doi=10.1086/667694|s2cid=84442763}}</ref> Among primates, this marked of a response to environmental stressors ([[phenotypic plasticity]]) is only demonstrated in modern humans.<ref name=":0"/><ref>{{Cite journal |last1=Antón |first1=Susan C. |last2=Potts |first2=Richard |last3=Aiello |first3=Leslie C. |date=2014-07-04 |title=Evolution of early ''Homo'': An integrated biological perspective |url=https://www.science.org/doi/10.1126/science.1236828 |journal=Science |language=en |volume=345 |issue=6192 |pages=1236828 |doi=10.1126/science.1236828 |pmid=24994657 |s2cid=30188239 |issn=0036-8075}}</ref><ref name=":02">{{Cite journal |last1=Antón |first1=Susan C. |last2=Taboada |first2=Hannah G. |last3=Middleton |first3=Emily R. |last4=Rainwater |first4=Christopher W. |last5=Taylor |first5=Andrea B. |last6=Turner |first6=Trudy R. |author-link6=Trudy Turner|last7=Turnquist |first7=Jean E. |last8=Weinstein |first8=Karen J. |last9=Williams |first9=Scott A. |date=2016-07-05 |title=Morphological variation in ''Homo erectus'' and the origins of developmental plasticity |journal=Philosophical Transactions of the Royal Society B: Biological Sciences |volume=371 |issue=1698 |pages=20150236 |doi=10.1098/rstb.2015.0236 |issn=0962-8436 |pmc=4920293 |pmid=27298467}}</ref> |
|||
Like modern humans and unlike other [[great ape]]s, there does not seem to have been a great size disparity between ''H. erectus'' males and females (size-specific [[sexual dimorphism]]), though there is not much fossil data regarding this.<ref name=Simpson2008/> Brain size in two adults from [[Koobi Fora]] measured {{cvt|848|and|804|cc}},<ref name=Anton2016>{{cite journal | vauthors = Antón SC, Taboada HG, Middleton ER, Rainwater CW, Taylor AB, Turner TR, Turnquist JE, Weinstein KJ, Williams SA | display-authors = 6 | title = Morphological variation in Homo erectus and the origins of developmental plasticity | journal = Philosophical Transactions of the Royal Society of London. Series B, Biological Sciences | volume = 371 | issue = 1698 | page = 20150236 | date = July 2016 | pmid = 27298467 | pmc = 4920293 | doi = 10.1098/rstb.2015.0236 }}</ref> and another significantly smaller adult measured {{cvt|691|cc}}, which could possibly indicate sexual dimorphism, though sex was undetermined.<ref name=Spoor2007>{{cite journal | vauthors = Spoor F, Leakey MG, Gathogo PN, Brown FH, Antón SC, McDougall I, Kiarie C, Manthi FK, Leakey LN | display-authors = 6 | title = Implications of new early Homo fossils from Ileret, east of Lake Turkana, Kenya | journal = Nature | volume = 448 | issue = 7154 | pages = 688–691 | date = August 2007 | pmid = 17687323 | doi = 10.1038/nature05986 | author2-link = Mary Leakey | s2cid = 35845 | bibcode = 2007Natur.448..688S }}</ref> Another case that depicts the difficulty of assigning sex to the fossil record is a few samples taken in Olduvai Gorge. In 1960, in Olduvai Gorge two skulls identified as OH12 and OH9, were found to be that of ''H. erectus'' with a cranial capacities of 1000 cc and 700 cc.<ref name=":5">{{Cite book |last=Leakey |first=Mary D |url=http://catalog.hathitrust.org/enwiki/api/volumes/oclc/6131892.html |title=Olduvai Gorge: my search for early man |date=1979 |publisher=Collins |location=London |isbn=9780002116138 |language=English |oclc=647137093}}</ref> It is unclear if sexual dimorphism is at play here since the remains are fragmentary.<ref name=":5" /> If ''H. erectus'' did not exhibit sexual dimorphism, then it is possible that they were the first in the human line to do so, though the fragmentary fossil record for earlier species makes this unclear. If yes, then there was a substantial and sudden increase in female height.<ref name=Plavcan2012>{{cite journal| vauthors = Plavcan JM |year=2012|title=Body Size, Size Variation, and Sexual Size Dimorphism in Early ''Homo''|journal=Current Anthropology|volume=53|issue=S6|pages=309–423|doi=10.1086/667605|s2cid=84095311}}</ref> Certain features of sexual dimorphism are often identified in the possibility of determining sex such as lack of muscle marking.<ref name=":13">{{Cite web |title=Rightmire GP. The Evolution of Homo Erectus: Comparative Anatomical Studies of an Extinct Human Species. Cambridge University Press; 1990. |url=https://search.library.ucr.edu/permalink/01CDL_RIV_INST/14qc2ti/alma991029515979704706 |access-date=2022-05-05 |website=search.library.ucr.edu |language=en}}</ref> |
|||
Springer (''see'' upper graph-model) depicts the presence of ''H. erectus'' as dominating the temporal and geographic development of human evolution; and as persisting broadly throughout Africa and Eurasia for nearly 2 million years, eventually evolving into ''[[Homo heidelbergensis|H. heidelbergensis / H. rhodesiensis]]'', which in turn evolved in ''H. sapiens''. Reed, et al. shows ''Homo ergaster'' as the ancestor of ''Homo erectus''; then it is ''ergaster'', or a variety of ''ergaster'', or perhaps a hybrid of ''ergaster'' and ''erectus'', which develops into species that evolve into archaic and then modern humans and then out of Africa. |
|||
[[File:Homo.erectus.adult.female.smithsonian.timevanson.flickr.jpg|thumb|Reconstruction of a female ''H. erectus'']] |
|||
''H. erectus'' had about the same limb configurations and proportions as modern humans, implying humanlike locomotion,<ref>{{cite journal | vauthors = Ruff C | title = Femoral/humeral strength in early African Homo erectus | journal = Journal of Human Evolution | volume = 54 | issue = 3 | pages = 383–390 | date = March 2008 | pmid = 17977577 | doi = 10.1016/j.jhevol.2007.09.001 | bibcode = 2008JHumE..54..383R }}</ref> the first in the ''Homo'' lineage.<ref name=":0">{{Cite journal |last1=Antón |first1=Susan C. |last2=Taboada |first2=Hannah G. |last3=Middleton |first3=Emily R. |last4=Rainwater |first4=Christopher W. |last5=Taylor |first5=Andrea B. |last6=Turner |first6=Trudy R. |last7=Turnquist |first7=Jean E. |last8=Weinstein |first8=Karen J. |last9=Williams |first9=Scott A. |date=2016-07-05 |title=Morphological variation in Homo erectus and the origins of developmental plasticity |journal=Philosophical Transactions of the Royal Society B: Biological Sciences |volume=371 |issue=1698 |pages=20150236 |doi=10.1098/rstb.2015.0236 |issn=0962-8436 |pmc=4920293 |pmid=27298467}}</ref> ''H. erectus'' tracks near [[Ileret]], Kenya, also indicate a [[human gait]].<ref name=Hatala/> A humanlike shoulder suggests an ability for high speed throwing.<ref name=":4">Roach, & Richmond. (2015). "Clavicle length, throwing performance and the reconstruction of the Homo erectus shoulder". ''Journal of Human Evolution'', 80(C), 107–113.</ref> It was once thought that Turkana boy had 6 [[lumbar vertebra]] instead of the 5 seen in modern humans and 11 instead of 12 [[thoracic vertebra]]e, but this has since been revised, and the specimen is now considered to have exhibited a humanlike curvature of the spine ([[lordosis]]) and the same number of respective vertebrae.<ref>{{cite journal | vauthors = Haeusler M, Schiess R, Boeni T | title = New vertebral and rib material point to modern bauplan of the Nariokotome Homo erectus skeleton | journal = Journal of Human Evolution | volume = 61 | issue = 5 | pages = 575–582 | date = November 2011 | pmid = 21868059 | doi = 10.1016/j.jhevol.2011.07.004 | bibcode = 2011JHumE..61..575H | url = https://www.zora.uzh.ch/id/eprint/50126/6/Haeusler_New_vertebral_and_rib_material_point_to_modern_bauplan.pdf }}</ref> |
|||
It is largely unclear when human ancestors lost most of their body hair. Genetic analysis suggests that high activity in the [[melanocortin 1 receptor]], which would produce dark skin, dates back to 1.2 Mya. This could indicate the evolution of hairlessness around this time, as a lack of body hair would have left the skin exposed to harmful [[UV radiation]].<ref>{{cite journal| vauthors = Rogers AR, Iltis D, Wooding S |year=2004|title=Genetic Variation at the MC1R Locus and the Time since Loss of Human Body Hair|journal=Current Anthropology|volume=45|issue=1|pages=105–108|doi=10.1086/381006|s2cid=224795768}}</ref> It is possible that exposed skin only became maladaptive in the Pleistocene, because the increasing [[axial tilt|tilt]] of the Earth (which also caused the [[Quaternary glaciation|ice ages]]) would have increased solar radiation bombardment- which would suggest that hairlessness first emerged in the australopithecines.<ref name=Gilligan2010/> However, australopithecines seem to have lived at much higher, much colder elevations—typically {{cvt|1000–1600|m}} where the nighttime temperature can drop to {{cvt|10|or|5|C|F}}—so they may have required hair to stay warm, unlike early ''Homo'' which inhabited lower, hotter elevations.<ref>{{cite journal | vauthors = Dávid-Barrett T, Dunbar RI | title = Bipedality and hair loss in human evolution revisited: The impact of altitude and activity scheduling | journal = Journal of Human Evolution | volume = 94 | pages = 72–82 | date = May 2016 | pmid = 27178459 | pmc = 4874949 | doi = 10.1016/j.jhevol.2016.02.006 | bibcode = 2016JHumE..94...72D }}</ref> Populations in higher latitudes potentially developed lighter skin to prevent [[vitamin D deficiency]].<ref>{{cite journal | vauthors = Jablonski NG | title = Human skin pigmentation as an example of adaptive evolution | journal = Proceedings of the American Philosophical Society | volume = 156 | issue = 1 | pages = 45–57 | date = March 2012 | pmid = 23035389 | jstor = 23558077 }}</ref> A 500–300 kya ''H. erectus'' specimen from Turkey was diagnosed with the earliest known case of [[tuberculous meningitis]], which is typically exacerbated in dark-skinned people living in higher latitudes due to vitamin D deficiency.<ref name="First Homo erectus from Turkey and">{{cite journal | vauthors = Kappelman J, Alçiçek MC, Kazanci N, Schultz M, Ozkul M, Sen S | title = First Homo erectus from Turkey and implications for migrations into temperate Eurasia | journal = American Journal of Physical Anthropology | volume = 135 | issue = 1 | pages = 110–116 | date = January 2008 | pmid = 18067194 | doi = 10.1002/ajpa.20739 }}</ref> Hairlessness is generally thought to have facilitated sweating,<ref>{{cite journal | vauthors = Best A, Kamilar JM | title = The evolution of eccrine sweat glands in human and nonhuman primates | journal = Journal of Human Evolution | volume = 117 | pages = 33–43 | date = April 2018 | pmid = 29544622 | doi = 10.1016/j.jhevol.2017.12.003 | bibcode = 2018JHumE.117...33B | s2cid = 3921318 }}</ref> but reduction of parasite load and [[sexual selection]] have also been proposed.<ref>{{cite book| vauthors = Pagel M, Bodmer W |year=2004|chapter=The Evolution of Human Hairlessness: Cultural Adaptations and the Ectoparasite Hypothesis|title=Evolutionary Theory and Processes: Modern Horizons|pages=329–335|publisher=Springer, Dordrecht|doi=10.1007/978-94-017-0443-4_17|isbn=978-94-017-0443-4}}</ref><ref>{{cite journal| vauthors = Gile J |year=2010|title=Naked Love: The Evolution of Human Hairlessness|journal=Biological Theory|volume=5|issue=4|pages=326–336|doi=10.1162/BIOT_a_00062|s2cid=84164968}}</ref> |
|||
Both models show the Asian variety of ''Homo erectus'' going extinct recently. And both models indicate species [[Archaic human admixture with modern humans|admixture]]: early modern humans spread from Africa across different regions of the globe and interbred with earlier descendants of ''[[H. heidelbergensis|H. heidelbergensis / H. rhodesiensis]]'', namely the Neanderthals, Denisovans, as well as unknown archaic African hominins. ''See'' [[Archaic human admixture with modern humans|admixture]]; and ''see'' [[Neanderthal admixture theory]].<ref name="Whitfield, John">{{cite web |url= http://www.sciam.com/article.cfm?id=lovers-not-fighters |title= Lovers not fighters|work= Scientific American |author= Whitfield, John |date= 18 February 2008}}</ref> |
|||
===Metabolism=== |
|||
==Use of tools and fire== |
|||
[[File:Pithecanthropus modjokertensis Tjokro Handojo.JPG|thumb|left|Front view of the [[Mojokerto child]] skull]] |
|||
The [[Paleolithic]] Age (Old Stone Age) of prehistoric [[human history]]<!------Yes, it's an oxymoron; but it's ok: pls see the linked article "History of the World"-----> and industry is dated from 2.6 million years ago to about 10,000 years ago;<ref name=Thoth&Schick>Toth, Nicholas; Schick, Kathy (2007). {{cite web|url=http://www.springerlink.com/content/u68378621542472j.html|title=Handbook of Paleoanthropology}} In Henke, H.C. Winfried; Hardt, Thorolf; Tatersall, Ian. ''Handbook of Paleoanthropology''. Volume 3. Berlin; Heidelberg; New York: Springer-Verlag. p. 1944. (PRINT: ISBN 978-3-540-32474-4 ONLINE: ISBN 978-3-540-33761-4)</ref> thus it closely coincides with the [[Pleistocene]] epoch of geologic time, which is 2.58 million to 11,700 years ago.<ref>{{cite web|url=http://www.ucmp.berkeley.edu/quaternary/pleistocene.php|title=The Pleistocene Epoch|publisher=University of California Museum of Paleontology|accessdate=22 August 2014}}</ref> The beginning of early human evolution reaches back to the earliest innovations of primitive technology and tool culture. |
|||
The 1.8 Ma [[Mojokerto child]] specimen from Java, who died at about 1 year of age, presented 72–84% of the average adult brain size, which is more similar to the faster brain growth trajectory of great apes than modern humans. This indicates that ''H. erectus'' was probably not cognitively comparable to modern humans, and that [[altriciality|secondary altriciality]]—an extended childhood and long period of dependency due to the great amount of time required for brain maturation—evolved much later in human evolution, perhaps in the modern human/Neanderthal last common ancestor.<ref name=Coqueugniot2004>{{cite journal | vauthors = Coqueugniot H, Hublin JJ, Veillon F, Houët F, Jacob T | title = Early brain growth in Homo erectus and implications for cognitive ability | journal = Nature | volume = 431 | issue = 7006 | pages = 299–302 | date = September 2004 | pmid = 15372030 | doi = 10.1038/nature02852 | s2cid = 4428043 | bibcode = 2004Natur.431..299C }}</ref> It was previously believed that, based on the narrow pelvis of Turkana boy, ''H. erectus'' could only safely deliver a baby with a brain volume of about {{cvt|230|cc}}, equating to a similar brain growth rate as modern humans to achieve the average adult brain size of {{cvt|600–1067|cc}}. However, a 1.8 Ma female pelvis from Gona<!--[[Gona]] directs to a village in Papua New Guinea-->, Ethiopia, shows that ''H. erectus'' babies with a brain volume of {{cvt|310|cc}} could have been safely delivered, which is 34–36% the mean adult size, compared to 40% in chimps and 28% in modern humans. This more aligns with the conclusions drawn from the Mojokerto child.<ref name=Simpson2008>{{cite journal | vauthors = Simpson SW, Quade J, Levin NE, Butler R, Dupont-Nivet G, Everett M, Semaw S | title = A female Homo erectus pelvis from Gona, Ethiopia | journal = Science | volume = 322 | issue = 5904 | pages = 1089–1092 | date = November 2008 | pmid = 19008443 | doi = 10.1126/science.1163592 | s2cid = 22191315 | bibcode = 2008Sci...322.1089S }}</ref> A faster development rate could indicate a lower expected lifespan.<ref>{{cite journal | vauthors = Caspari R, Lee SH | title = Older age becomes common late in human evolution | journal = Proceedings of the National Academy of Sciences of the United States of America | volume = 101 | issue = 30 | pages = 10895–10900 | date = July 2004 | pmid = 15252198 | pmc = 503716 | doi = 10.1073/pnas.0402857101 | doi-access = free }}</ref> |
|||
Based on an average mass of {{cvt|63|kg}} for males and {{cvt|52.3|kg}} for females, the daily energy expenditure (DEE)—the amount of calories metabolized in one day—was estimated to be about 2271.8 and 1909.5 [[kcal]], respectively. This is similar to that of earlier ''Homo'', despite a marked increase in activity and migratory capacity, likely because the longer legs of ''H. erectus'' were more energy-efficient in long-distance movement. Nonetheless, the estimate for ''H. erectus'' females is 84% higher than that for ''Australopithecus'' females, possibly due to an increased body size and a decreased growth rate.<ref>{{cite journal | vauthors = Steudel-Numbers KL | title = Energetics in Homo erectus and other early hominins: the consequences of increased lower-limb length | journal = Journal of Human Evolution | volume = 51 | issue = 5 | pages = 445–453 | date = November 2006 | pmid = 16780923 | doi = 10.1016/j.jhevol.2006.05.001 | bibcode = 2006JHumE..51..445S }}</ref> A 2011 study, assuming high energy or dietary fat requirements based on the abundance of large game animals at ''H. erectus'' sites, calculated a DEE of 2,700–3,400 kcal of which 27–44% derived from fat, and 44–62% of the fat from animal sources. In comparison, modern humans with a similar activity level have a DEE of 2,450 calories, of which 33% derives from fat, and 49% of the fat from animals.<ref name=BenDor2011/> |
|||
''[[Homo ergaster]] '' used more diverse and sophisticated [[stone tool]]s than its predecessors, where early ''Homo erectus'' used comparatively primitive tools. This is probably because ''H. ergaster'' inherited, used, and created tools first of [[Oldowan]] technology and later advanced the technology to the [[Acheulean]].<ref>{{cite book | author = Beck, Roger B.; Black, Linda; Krieger, Larry S.; Naylor, Phillip C. and Shabaka, Dahia Ibo | title = World History: Patterns of Interaction | publisher = McDougal Littell | year = 1999 | location = Evanston, IL |isbn = 0-395-87274-X }}</ref> Because the use of Acheulean tools began ca. 1.8 million years ago,<ref>The Earth Institute. (2011-09-01). [http://www.earth.columbia.edu/articles/view/2839 Humans Shaped Stone Axes 1.8 Million Years Ago, Study Says]. Columbia University. Accessed 5 January 2012.</ref> and the line of ''H. erectus'' diverged some 200,000 years before the general innovation of Acheulean industry in Africa, then it is plausible that the Asian migratory descendants of ''H. erectus'' made no use of Acheulean technology. It has been suggested that the Asian ''H. erectus'' may have been the first humans to use rafts to travel over bodies of water, including oceans.<ref>{{cite journal | title = Paleoanthropology: Ancient Island Tools Suggest Homo erectus Was a Seafarer | journal = Science | volume = 279 | issue = 5357 | pages = 1635–1637 | date = 13 March 1998 | author = Gibbons, Ann | doi = 10.1126/science.279.5357.1635}}</ref> And the oldest stone tool found in [[Turkey]] reveals that hominins passed through the [[Anatolia]]n gateway from western Asia to Europe approximately 1.2 million years ago—much earlier than previously thought.<ref>[http://www.sciencedaily.com/releases/2014/12/141223084139.htm Oldest stone tool ever found in Turkey discovered] by the University of Royal Holloway London and published in ScienceDaily on December 23, 2014</ref> |
|||
=== |
===Bone thickness=== |
||
[[File:Homo erectus humeri.png|thumb|Cross sections of Chinese ''H. erectus'' [[humerus|humeri]] (upper arm bones) showing extremely thickened [[cortical bone]]]] |
|||
[[East Africa]]n sites, such as [[Chesowanja]] near [[Lake Baringo]], [[Koobi Fora]], and [[Olorgesailie]] in [[Kenya]], show potential evidence that fire was utilized by early humans. At Chesowanja, archaeologists found fire-hardened clay fragments, dated to 1.42 mya.<ref name="James">{{cite journal|last=James|first=Steven R.|date=February 1989|title=Hominid Use of Fire in the Lower and Middle Pleistocene: A Review of the Evidence|journal=Current Anthropology |url=http://faculty.ksu.edu.sa/archaeology/Publications/Hearths/Hominid%20Use%20of%20Fire%20in%20the%20Lower%20and%20Middle%20Pleistocene.pdf |volume=30|issue=1|pages=1–26|publisher=University of Chicago Press|doi=10.1086/203705 |accessdate=2012-04-04}}</ref> Analysis showed that, in order to harden it, the clay must have been heated to about {{convert|400|C}}. At Koobi Fora, two sites show evidence of control of fire by ''Homo erectus'' at about 1.5 mya, with reddening of sediment associated with heating the material to {{convert|200|-|400|C|F|abbr=on}}.<ref name="James" /> At a "hearth-like depression" at a site in Olorgesailie, Kenya, some microscopic [[charcoal]] was found—but that could have resulted from natural brush fires.<ref name="James"/> |
|||
The [[cortical bone]] (the outer layer of the bone) is extraordinarily thickened, particularly in East Asian populations. The skullcaps have oftentimes been confused with fossil turtle [[carapace]]s,<ref name=Boaz2004/> and the [[medullary canal]] in the [[long bone]]s (where the [[bone marrow]] is stored, in the limbs) is extremely narrowed (medullary [[stenosis]]). This degree of thickening is usually exhibited in semi-aquatic animals which used their heavy ([[pachyosteosclerosis|pachyosteosclerotic]]) bones as ballasts to help them sink, induced by [[hypothyroidism]]. Male specimens have thicker cortical bone than females.<ref name=Kennedy1985/> |
|||
It is largely unclear what function this could have served. All pathological inducers would leave scarring or some other indicator not normally exhibited in ''H. erectus''. Before more complete skeletons were discovered, Weidenreich suggested ''H. erectus'' was a gigantic species, thickened bone required to support the massive weight. It was hypothesized that intense physical activity could have induced bone thickening, but in 1970, human biologist [[Stanley Marion Garn]] demonstrated there is a low correlation between the two at least in modern humans. Garn instead noted different races have different average cortical bone thicknesses, and concluded it is genetic rather than environmental. It is unclear if the condition is caused by increased bone apposition (bone formation) or decreased [[bone resorption]], but Garn noted the stenosis is quite similar to the [[congenital]] condition in modern humans induced by hyper-apposition. In 1985, biological anthropologist Gail Kennedy argued for resorption as a result of [[hyperparathyroidism]] caused by [[hypocalcemia]] ([[calcium in biology|calcium]] deficiency), a consequence of a dietary shift to low-calcium meat. Kennedy could not explain why the [[calcium metabolism]] of ''H. erectus'' never adjusted.<ref name=Kennedy1985>{{cite journal | vauthors = Kennedy GE |year=1985|title=Bone thickness in ''Homo erectus''|journal=Journal of Human Evolution|volume=14|issue=8|pages=699–708|doi=10.1016/S0047-2484(85)80052-X|bibcode=1985JHumE..14..699K }}</ref> In 1985, American paleoanthropologist Mary Doria Russell<!--not the novelist--> and colleagues argued the supraorbital torus is a response to withstanding major [[bending moment]] which localizes in that region when significant force is applied through the front teeth, such as while using the mouth as a third hand to carry objects.<ref>{{cite journal| vauthors = Russell MD, Brown T, Garn SM, Giris F, Turkel S, İşcan MY, Oyen OJ, Jacobshagen B, Pietrusewsky M, Rightmire GP, Smith FH | display-authors = 6 |year=1985|title=The Supraorbital Torus: 'A Most Remarkable Peculiarity'|journal=Current Anthropology|volume=26|issue=3|pages=337–350|doi=10.1086/203279|s2cid=146857927}}</ref> |
|||
In [[Gadeb]], [[Ethiopia]], fragments of [[Tuff#Welded tuff|welded tuff]] that appeared to have been burned, or scorched, were found alongside ''H. erectus''–created [[Acheulean]] artifacts; but such re-firing of the rocks may have been caused by local volcanic activity.<ref name="James"/> In the [[Middle Awash]] River Valley, cone-shaped depressions of reddish clay were found that could have been created only by temperatures of {{convert|200|C|abbr=on}} or greater. These features are thought to be burnt tree stumps such that the fire was likely away from a habitation site.<ref name="James"/> Burnt stones are found in the Awash Valley, but naturally burnt (volcanic) welded tuff is also found in the area. |
|||
In 2004, Noel Boaz and Russel Ciochon suggested it was a result of a cultural practice, wherein ''H. erectus'' would fight each other with fists, stones, or clubs to settle disputes or battle for mates, since the skull is reinforced in key areas. The mandible is quite robust, capable of absorbing heavy blows (no "glass jaw"); the heavy brow ridge protects the eyes, and transitions into a bar covering the ears, connecting all the way in the back of the skull, meaning blows to any of these regions can be effectively dissipated across the skull; and the sagittal keel protects the top of the braincase. Many skullcaps bear usually debilitating fractures, such as the Peking Man skull X, yet they can show signs of surviving and healing. Anthropologist [[Peter Brown (anthropologist)|Peter Brown]] suggested a similar reason for the unusual thickening of the modern [[Australian Aboriginal]] skull, a result of a ritual popular in central and southeast Australian tribes where adversaries would wack each other with [[waddy|waddies]] (sticks) until [[knockout]].<ref name=Boaz2004>{{cite journal| vauthors = Boaz N, Ciochon R |year=2004|title=Headstrong Hominids|url=https://www.researchgate.net/publication/295215638|journal=Natural History|volume=113|issue=1|pages=28–34}}</ref> |
|||
A site at [[Bnot Ya'akov Bridge]], [[Israel]] is reported to evidence that ''H. erectus'' or ''H. ergaster'' controlled fire there between 790,000 and 690,000 BP;<ref name="Rincon">{{cite news|first=Paul|last=Rincon|url=http://news.bbc.co.uk/2/hi/science/nature/3670017.stm | title=Early human fire skills revealed|publisher=[[BBC News]]|date=29 April 2004|accessdate=2007-11-12}}</ref> to date this claim has been widely accepted. Some evidence is found that ''H. erectus'' was controlling fire less than 250,000 years ago. Also, that ''H. erectus'' was cooking their food as early as 500,000 years ago.<ref>{{Cite book|title = Worlds Together, Worlds Apart|last = Pollard|first = Elizabeth|publisher = Norton|year = 2015|isbn = 978-0-393-92207-3|location = New York|pages = 13}}</ref> Re-analysis of burnt bone fragments and plant ashes from the [[Wonderwerk Cave]], South Africa, has been dubbed evidence supporting human control of fire there by 1 mya.<ref name=Pringle2012>{{citation|date=2 April 2012 |author=Pringle, Heather |title=Quest for Fire Began Earlier Than Thought |journal=ScienceNOW |publisher=[[American Association for the Advancement of Science]] |url=http://news.sciencemag.org/sciencenow/2012/04/quest-for-fire-began-earlier-tha.html?ref=em |accessdate=2012-04-04}}</ref> |
|||
== |
==Culture== |
||
===Social structure=== |
|||
There is no archaeological evidence that ''Homo erectus'' cooked their food. The idea has been suggested,<ref>{{cite book|last=Wrangham|first=Richard|title=Catching Fire|year=2009|publisher=Basic Books}}</ref> but is not generally accepted.<ref>{{cite book|title=Female Hierarchies|year=1972|publisher=Beresford Book Service|pages=220–229|author=Adrienne Zihlman|author2=Nancy Tanner|editor=Lionel Tiger, Heather T. Fowler|chapter=Gathering and the Hominid Adaptation}}</ref><ref>{{cite journal|last=Fedigan|first=Linda Marie|title=The Changing Role of Women in Models of Human Evolution|journal=Annual Review of Anthropology|year=1986|volume=15|pages=25–66|doi=10.1146/annurev.an.15.100186.000325}}</ref> It is known, from the study of [[Use-wear analysis|use-wear]] on [[handaxes]], that meat formed a growing part of the ''H. erectus'' diet. But meat is digestible without cooking, and its use is not itself evidence of cooking. As nuts, berries, fruits, and grasses are also eaten raw, then cooking cannot be presumed: the issue rests on clear evidence from archaeological sites, which at present does not exist.{{or|date=August 2015}} |
|||
[[File:Ileret trackways.jpg|thumb|upright=1.3|Diagram of fossil trackways from 2 sites near [[Ileret]], Kenya]] |
|||
The only fossil evidence regarding ''H. erectus'' group composition comes from four sites outside of [[Ileret]], Kenya, where 97 footprints made 1.5 Mya were likely left by a group of at least 20 individuals. One of these trackways, based on the size of the footprints, may have been an entirely male group, which could indicate they were some specialised task group, such as a hunting or foraging party, or a border patrol. If correct, this would also indicate sexual division of labour, which distinguishes human societies from those of other great apes and social mammalian carnivores. In modern hunter gatherer societies who target large prey items, typically male parties are dispatched to bring down these high-risk animals, and, due to the low success rate, female parties focus on more predictable foods.<ref name=Hatala>{{cite journal | vauthors = Hatala KG, Roach NT, Ostrofsky KR, Wunderlich RE, Dingwall HL, Villmoare BA, Green DJ, Harris JW, Braun DR, Richmond BG | display-authors = 6 | title = Footprints reveal direct evidence of group behavior and locomotion in Homo erectus | journal = Scientific Reports | volume = 6 | issue = 28766 | pages = 28766 | date = July 2016 | pmid = 27403790 | pmc = 4941528 | doi = 10.1038/srep28766 | doi-access = free | bibcode = 2016NatSR...628766H }}</ref> Based on modern day savanna chimp and [[baboon]] group composition and behavior, ''H. erectus ergaster'' may have lived in large, multi-male groups in order to defend against large savanna predators in the open and exposed environment.<ref name=Willems2017/> However, dispersal patterns indicate that ''H. erectus'' generally avoided areas with high carnivore density.<ref name=Carotenuto2016/> It is possible that male–male bonding and male–female friendships were important societal aspects.<ref name=Willems2017>{{cite journal | vauthors = Willems EP, van Schaik CP | title = The social organization of Homo ergaster: Inferences from anti-predator responses in extant primates | journal = Journal of Human Evolution | volume = 109 | pages = 11–21 | date = August 2017 | pmid = 28688456 | doi = 10.1016/j.jhevol.2017.05.003 | bibcode = 2017JHumE.109...11W }}</ref> |
|||
Because ''H. erectus'' children had faster brain growth rates, ''H. erectus'' likely did not exhibit the same degree of maternal investment or child-rearing behaviours as modern humans.<ref name=Simpson2008/> |
|||
==Sociality== |
|||
''Homo erectus'' was probably the first [[hominin]] to live in a [[hunter-gatherer]] society, and anthropologists such as [[Richard Leakey]] believe that ''erectus'' was socially more like modern humans than the more ''[[Australopithecus]]''-like species before it. Likewise, increased cranial capacity generally coincides with the more sophisticated tools occasionally found with fossils. |
|||
Because ''H. erectus'' males and females are thought to have been about the same size compared to other great apes (exhibit less size-specific sexual dimorphism), it is generally hypothesised that they lived in a monogamous society, as reduced sexual dimorphism in primates is typically correlated with this mating system.<ref name=Plavcan2012/> However, it is unclear if ''H. erectus'' did in fact exhibit humanlike rates of sexual dimorphism.<ref name=Spoor2007/> If they did, then it would mean only female height increased from the ancestor species, which could have been caused by a shift in female fertility or diet, and/or reduced pressure on males for large size. This in turn could imply a shift in female behavior which made it difficult for males to maintain a harem, and vice versa.<ref>{{cite journal | vauthors = Plavcan JM |year=2012|title=Implications of Male and Female Contributions to Sexual Size Dimorphism for Inferring Behavior in the Hominin Fossil Record|journal=International Journal of Primatology|volume=33|issue=6|pages=1364–1381|doi=10.1007/s10764-012-9642-z|s2cid=17850676}}</ref> |
|||
The discovery of [[Turkana boy]] (''H. ergaster'') in 1984 evidenced that, despite its ''Homo sapiens''-like anatomy, ''ergaster'' may not have been capable of producing sounds comparable to modern human [[Speech communication|speech]]. It likely communicated in a [[Origin of language#Early Homo|proto-language]] lacking the fully developed structure of modern human language but more developed than the non-verbal communication used by [[chimpanzee]]s.<ref>{{cite book |author=Ruhlen, Merritt|title=The origin of language: tracing the evolution of the mother tongue |publisher=Wiley |location=New York |year=1994 |isbn=0-471-58426-6}}</ref> This inference is challenged by the find in [[Homo erectus#Homo erectus georgicus|Dmanisi]], Georgia, of an ''H. ergaster'' / ''erectus'' [[vertebra]]e (at least 150,000 years earlier than the Turkana Boy) that reflects vocal capabilities within the range of ''H. sapiens''.<ref name="Bower, Bruce 275–276"/> Both brain size and the presence of the [[Broca's area]] also support the use of articulate language.<ref>{{cite book|title=Origins Reconsidered|author=Richard Leakey|year=1992|publisher=Anchor|pages=257–258|isbn=0-385-41264-9}}</ref> |
|||
===Food=== |
|||
''H. erectus'' was probably the first hominin to live in small, familiar [[band societies|band-societies]] similar to modern hunter-gatherer band-societies;<ref>{{cite book |author=Boehm, Christopher |title=Hierarchy in the forest: the evolution of egalitarian behavior |publisher=Harvard University Press |location=Cambridge |year=1999|isbn=0-674-39031-8|url=http://books.google.com/books?id=ljxS8gUlgqgC&pg=PA198 |page=198}}</ref> and is thought to be the first hominin species to hunt in coordinated groups, to use complex tools, and to care for infirm or weak companions. |
|||
Increasing brain size is often directly associated with a meatier diet and resultant higher caloric intake. [[Human entomophagy]] and therefore an increase in protein consumption through insects has also been proposed as a possible cause. However, it is also possible that the energy-expensive guts decreased in size in ''H. erectus'', because the large ape gut is used to synthesize fat by fermenting plant matter, which was replaced by dietary animal fat, allowing more energy to be diverted to brain growth. This would have increased brain size indirectly while maintaining the same caloric requirements of ancestor species. ''H. erectus'' may have also been the first to use a [[Hunter-gatherer|hunting and gathering]] food collecting strategy as a response to the increasing dependence on meat. With an emphasis on teamwork, division of labor, and food sharing, hunting and gathering was a dramatically different subsistence strategy from previous modes.<ref name=Ungar2006/><ref name=BenDor2011/> |
|||
[[File:Reproducción de Palaeoloxodon antiquus.jpg|thumb|upright=1|''H. erectus'' ate primarily large game, such as the [[straight-tusked elephant]] (above)]] |
|||
There has been debate as to whether ''H. erectus'',<ref name="Whitfield, John"/> and possibly the later [[Homo neanderthalensis|Neanderthal]]s,<ref>{{cite web |url= http://news.nationalgeographic.com/news/2006/10/061030-neanderthals.html |title= Neanderthals, Modern Humans Interbred, Bone Study Suggests |
|||
''H. erectus'' sites frequently are associated with assemblages of medium- to large-sized game, namely [[elephant]]s, [[rhino]]s, [[hippo]]s, [[bovine]]s, and [[boar]]s. ''H. erectus'' would have had considerable leftovers, potentially pointing to food sharing or long-term [[food preservation]] (such as by drying) if most of the kill was indeed utilized. It is possible that ''H. erectus'' grew to become quite dependent on large-animal meat, and the disappearance of ''H. erectus'' from the [[Levant]] is correlated with the local extinction of the [[straight-tusked elephant]].<ref name=BenDor2011>{{cite journal | vauthors = Ben-Dor M, Gopher A, Hershkovitz I, Barkai R | title = Man the fat hunter: the demise of Homo erectus and the emergence of a new hominin lineage in the Middle Pleistocene (ca. 400 kyr) Levant | journal = PLOS ONE | volume = 6 | issue = 12 | pages = e28689 | year = 2011 | pmid = 22174868 | pmc = 3235142 | doi = 10.1371/journal.pone.0028689 | bibcode = 2011PLoSO...628689B | doi-access = free }}</ref> Nonetheless, ''H. erectus'' diet likely varied widely depending upon location. For example, at the 780 kya [[Daughters of Jacob Bridge|Gesher Benot Ya'aqov]] site, Israel, the inhabitants gathered and ate 55 different types of fruits, vegetables, seeds, nuts, and tubers, and it appears that they used fire to roast certain plant materials that otherwise would have been inedible; they also consumed amphibians, reptiles, birds, aquatic and terrestrial invertebrates, in addition to the usual large creatures such as elephant and [[fallow deer]].<ref>{{cite journal | vauthors = Melamed Y, Kislev ME, Geffen E, Lev-Yadun S, Goren-Inbar N | title = The plant component of an Acheulian diet at Gesher Benot Ya'aqov, Israel | journal = Proceedings of the National Academy of Sciences of the United States of America | volume = 113 | issue = 51 | pages = 14674–14679 | date = December 2016 | pmid = 27930293 | pmc = 5187744 | doi = 10.1073/pnas.1607872113 | bibcode = 2016PNAS..11314674M | doi-access = free }}</ref> At the 1.95 Mya FwJJ20 lakeside site in the [[East Turkana]] Basin, Kenya, the inhabitants ate (alongside the usual bovids, hippos, and rhinos) aquatic creatures such as [[turtle]]s, [[crocodile]]s, and [[catfish]]. The large animals were likely scavenged at this site, but the turtles and fish were possibly collected live.<ref>{{cite journal | vauthors = Steele TE | title = A unique hominin menu dated to 1.95 million years ago | journal = [[Proceedings of the National Academy of Sciences of the United States of America]] | volume = 107 | issue = 24 | pages = 10771–10772 | date = June 2010 | pmid = 20534542 | pmc = 2890732 | doi = 10.1073/pnas.1005992107 | doi-access = free | bibcode = 2010PNAS..10710771S }}</ref> In East Africa between 2.0 and 1.4 Mya, carcasses of {{C4}}-grazing ungulates, particularly [[Alcelaphini|alcelaphins]], featured increasingly prominently in the diet of these hominins.<ref>{{cite journal |last1=Patterson |first1=D. B. |last2=Braun |first2=D. R. |last3=Behrensmeyer |first3=A. K. |last4=Merritt |first4=S. |last5=Zliobaite |first5=I. |last6=Reeves |first6=J. S. |last7=Wood |first7=B. A. |last8=Fortelius |first8=M. |last9=Bobe |first9=R. |date=1 September 2017 |title=Ecosystem evolution and hominin paleobiology at East Turkana, northern Kenya between 2.0 and 1.4 Ma |journal=[[Palaeogeography, Palaeoclimatology, Palaeoecology]] |volume=481 |pages=1–13 |doi=10.1016/j.palaeo.2017.05.001 |bibcode=2017PPP...481....1P |doi-access=free }}</ref> At the 1.5 Mya [[Trinil H. K. Fauna|Trinil H. K.]] site, Java, ''H. erectus'' likely gathered fish and shellfish.<ref>{{cite journal | vauthors = Joordens JC, Wesselingh FP, de Vos J, Vonhof HB, Kroon D | title = Relevance of aquatic environments for hominins: a case study from Trinil (Java, Indonesia) | journal = [[Journal of Human Evolution]] | volume = 57 | issue = 6 | pages = 656–671 | date = December 2009 | pmid = 19683789 | doi = 10.1016/j.jhevol.2009.06.003 | bibcode = 2009JHumE..57..656J }}</ref> |
|||
|work= National Geographic News |author= Owen, James |date=30 October 2006 |
|||
|accessdate=2008-01-14 }}</ref> may have interbred with [[anatomically modern human]]s in [[Europe]] and [[Asia]]. ''See'' [[Neanderthal admixture theory]]. |
|||
Dentally, ''H. erectus'' mouths were not as versatile as those of ancestor species, capable of processing a narrower range of foods. However, tools were likely used to process hard foods, thus affecting the chewing apparatus. This combination may have instead increased dietary flexibility (though this does not equate to a highly varied diet). Such versatility may have permitted ''H. erectus'' to inhabit a range of different environments and migrate beyond Africa.<ref name=Ungar2006/> |
|||
==Descendants and subspecies== |
|||
''Homo erectus'' is the most, or one of the most, long-lived species of ''Homo'', having existed well over one million years and perhaps over two million years; ''Homo sapiens'' has existed for about 200,000 years. If considering ''Homo erectus'' in its strict sense (that is, as referring to only the Asian variety) no consensus has been reached as to whether it is ancestral to ''H. sapiens'' or any later hominins (''see'' above, "Interpreting evolution: ..."). |
|||
[[File:Homo erectus adult female - head model - Smithsonian Museum of Natural History - 2012-05-17.jpg|right|thumb|A model of the face of an adult female ''Homo erectus''. Reconstruction by [[John Gurche]], [[Smithsonian Museum of Natural History]], based on [[KNM ER 3733]] and [[KNM ER 992|992]].]] |
|||
In 1999, British anthropologist [[Richard Wrangham]] proposed the "cooking hypothesis" which states that ''H. erectus'' speciated from the ancestral ''H. habilis'' because of fire usage and cooking 2 million years ago to explain the rapid doubling of brain size between these two species in only a 500,000-year timespan, and the sudden appearance of the typical human body plan. Cooking makes protein more easily digestible, speeds up nutrient absorption, and destroys food-borne pathogens, which would have increased the environment's natural carrying capacity, allowing group size to expand, causing selective pressure for sociality, requiring greater brain function.<ref name=Gowlett2016/><ref name=Gowlett2013/> However, the fossil record does not associate the emergence of ''H. erectus'' with fire usage nor with any technological breakthrough for that matter, and cooking likely did not become a common practice until after 400 kya.<ref name=Ungar2006/><ref name=BenDor2011/> |
|||
===''Homo erectus''=== |
|||
** ''[[Homo erectus erectus]]'' ([[Java Man]]) |
|||
** ''[[Homo erectus yuanmouensis]]'' ([[Yuanmou Man]]) |
|||
** ''[[Homo erectus lantianensis]]'' ([[Lantian Man]]) |
|||
** ''[[Homo erectus nankinensis]]'' ([[Nanjing Man]]) |
|||
** ''[[Homo erectus pekinensis]]'' ([[Peking Man]]) |
|||
** ''[[Homo erectus palaeojavanicus]]'' ([[Meganthropus]]) |
|||
** ''[[Homo erectus soloensis]]'' ([[Solo Man]]) |
|||
** ''[[Homo erectus tautavelensis]]'' ([[Tautavel Man]]) |
|||
** ''[[Homo erectus georgicus]]'' |
|||
Java Man's dispersal through Southeast Asia coincides with the [[extirpation]] of the giant turtle ''[[Megalochelys]]'', possibly due to overhunting as the turtle would have been an easy, slow-moving target which could have been stored for quite some time.<ref>{{Cite book|url=http://www.iucn-tftsg.org/cbftt/|title=Conservation Biology of Freshwater Turtles and Tortoises|chapter=Turtles and Tortoises of the World During the Rise and Global Spread of Humanity: First Checklist and Review of Extinct Pleistocene and Holocene Chelonians |year=2015|publisher=Chelonian Research Foundation|isbn=978-0-9653540-9-7| veditors = Rhodin A, Pritchard P, van Dijk PP, Saumure R, Buhlmann K, Iverson J, Mittermeier R |edition=First|series=Chelonian Research Monographs|volume=5|doi=10.3854/crm.5.000e.fossil.checklist.v1.2015|page=15 |last1=Rhodin |first1=Anders |last2=Thomson |first2=Scott |last3=Georgalis |first3=Georgios |last4=Karl |first4=Hans-Volker |last5=Danilov |first5=Igor |last6=Takahashi |first6=Akio |last7=de la Fuente |first7=Marcelo |last8=Bourque |first8=Jason |last9=Delfino |first9=Massimo |last10=Bour |first10=Roger |last11=Iverson |first11=John |last12=Shaffer |first12=Bradley |last13=Van Dijk |first13=Peter Paul }}</ref> |
|||
===Related species=== |
|||
* ''[[Homo ergaster]]'' |
|||
* ''[[Homo floresiensis]]'' |
|||
* ''[[Homo antecessor]]'' |
|||
* ''[[Homo heidelbergensis]]'' |
|||
* ''[[Human|Homo sapiens]]'' |
|||
** ''[[Homo sapiens idaltu]]'' |
|||
** ''[[Homo sapiens sapiens]]'' |
|||
* ''[[Neanderthal|Homo neanderthalensis]]'' |
|||
* ''[[Homo rhodesiensis]]'' |
|||
* ''[[Homo cepranensis]]'' |
|||
===Technology=== |
|||
===Previously referred taxa=== |
|||
====Tool production==== |
|||
* ''[[Wushan Man|Homo erectus wushanensis]]'' (actually a stem-[[orangutan]]) |
|||
{{Multiple image|image1=Canto tallado 2-Guelmim-Es Semara.jpg|caption1=[[Oldowan]] choppers did not become completely replaced until about 1 Mya|image2=Bifaz cordiforme.jpg|caption2=An [[Acheulean]] [[cordiform axe]]|direction=vertical}} |
|||
The discovery of ''[[Homo floresiensis]]'' in 2003 and of the recentness of its extinction has raised the possibility that numerous descendant species of ''Homo erectus'' may have existed in the islands of [[Southeast Asia]] and await fossil discovery (see ''[[Orang Pendek]]''). ''Homo erectus soloensis'', who was long assumed to have lived on Java at least as late as about 50,000 years ago but was re-dated in 2011 to a much higher age,<ref>[http://www.terradaily.com/reports/Finding_showing_human_ancestor_older_than_previously_thought_offers_new_insights_into_evolution_999.html Finding showing human ancestor older than previously thought offers new insights into evolution], 5 July 2011.</ref> would be one of them. Some scientists are skeptical of the claim that ''Homo floresiensis'' is a descendant of ''Homo erectus''. One explanation holds that the fossils are of a modern human with [[microcephaly]], while another one holds that they are from a group of [[pygmies]]. |
|||
''H. erectus'' is credited with inventing the [[Acheulean]] stone tool industry, succeeding the [[Oldowan]] industry,<ref>{{cite book | vauthors = Beck RB, Black L, Krieger LS, Naylor PC, Shabaka DI |title=World History: Patterns of Interaction |url=https://archive.org/details/mcdougallittellw00beck |url-access=registration |publisher=McDougal Littell |year=1999 |location=Evanston, IL |isbn=978-0-395-87274-1 }}{{page needed|date=December 2019}}</ref><ref>{{cite journal | vauthors = Richards MP | title = A brief review of the archaeological evidence for Palaeolithic and Neolithic subsistence | journal = European Journal of Clinical Nutrition | volume = 56 | issue = 12 | pages = 1270–1278 | date = December 2002 | pmid = 12494313 | doi = 10.1038/sj.ejcn.1601646 | doi-access = free }}</ref> and were the first to make [[lithic flake]]s bigger than {{cvt|10|cm}}, and [[hand axe]]s (which includes bifacial tools with only 2 sides, such as picks, knives, and [[cleaver (tool)|cleavers]]).<ref>{{cite journal | vauthors = de la Torre I | title = The origins of the Acheulean: past and present perspectives on a major transition in human evolution | journal = Philosophical Transactions of the Royal Society of London. Series B, Biological Sciences | volume = 371 | issue = 1698 | pages = 20150245 | date = July 2016 | pmid = 27298475 | pmc = 4920301 | doi = 10.1098/rstb.2015.0245 }}</ref> Though larger and heavier, these hand axes had sharper, chiseled edges.<ref name=Lepre2011/> They were likely multi-purpose tools, used in variety of activities such as cutting meat, wood, or edible plants.<ref name=Nowell2009/> In 1979, American paleontologist Thomas Wynn stated that Acheulean technology required operational intelligence (foresight and planning), being markedly more complex than Oldowan technology which included lithics of unstandardized shape, cross-sections, and symmetry. Based on this, he concluded that there is not a significant disparity in intelligence between ''H. erectus'' and modern humans and that, for the last 300,000 years, increasing intelligence has not been a major influencer of cultural evolution.<ref>{{cite journal| vauthors = Wynn T |year=1979|title=The Intelligence of Later Acheulean Hominids|journal=Man|volume=14|issue=3|pages=371–391|doi=10.2307/2801865|jstor=2801865}}</ref> However, a 1 year old ''H. erectus'' specimen shows that this species lacked an extended childhood required for greater brain development, indicating lower cognitive capabilities.<ref name=Coqueugniot2004/> A few sites, likely due to occupation over several generations, features hand axes en masse, such as at [[Melka Kunture]], Ethiopia; [[Olorgesailie]], Kenya; [[Isimila]], Tanzania; and [[Kalambo Falls]], Zambia.<ref name=Nowell2009/> |
|||
The earliest record of Acheulean technology comes from [[West Turkana]], Kenya 1.76 Mya. Oldowan lithics are also known from the site, and the two seemed to coexist for some time. The earliest records of Acheulean technology outside of Africa date to no older than 1 Mya, indicating it only became widespread after some secondary ''H. erectus'' dispersal from Africa.<ref name=Lepre2011>{{cite journal | vauthors = Lepre CJ, Roche H, Kent DV, Harmand S, Quinn RL, Brugal JP, Texier PJ, Lenoble A, Feibel CS | display-authors = 6 | title = An earlier origin for the Acheulian | journal = Nature | volume = 477 | issue = 7362 | pages = 82–85 | date = August 2011 | pmid = 21886161 | doi = 10.1038/nature10372 | s2cid = 4419567 | bibcode = 2011Natur.477...82L }}</ref> |
|||
==Individual fossils== |
|||
[[File:Pithecanthropus-erectus.jpg|thumb|Original fossils of ''Pithecanthropus erectus'' (now ''Homo erectus'') found in [[Java]] in 1891.]] |
|||
On Java, ''H. erectus'' produced tools from shells at [[Sangiran]]<ref>{{cite journal| vauthors = Choi K, Driwantoro D |year=2007|title=Shell tool use by early members of ''Homo erectus'' in Sangiran, central Java, Indonesia: cut mark evidence|journal=Journal of Archaeological Science|volume=34|issue=1|pages=48–58|doi=10.1016/j.jas.2006.03.013|bibcode=2007JArSc..34...48C }}</ref> and Trinil.<ref name=":1"/> Spherical stones, measuring {{cvt|6–12|cm}} in diameter, are frequently found in African and Chinese Lower Paleolithic sites, and were potentially used as [[bolas]]; if correct, this would indicate string and cordage technology.<ref>{{cite book|url={{google books|plainurl=yes|id=ODtqDQAAQBAJ|page=6}}| vauthors = Turner J |author-link1=Trudy Turner|year=1996|title=History and Science of Knots|publisher=World Scientific|pages=6–8|isbn=9789810224691}}</ref> |
|||
{{anchor|Fire}} |
|||
====Fire==== |
|||
{{See also|Control of fire by early humans}} |
|||
''H. erectus'' is credited as the first human ancestor to have used fire, though the timing of this invention is debated mainly because campfires very rarely and very poorly preserve over long periods of time, especially thousands or millions of years. The earliest claimed fire sites are in Kenya, FxJj20 at [[Koobi Fora]]<ref>{{cite journal| vauthors = Hlubik S, Berna F, Feibel C, Braun D |year=2017|title=Researching the Nature of Fire at 1.5 Mya on the Site of FxJj20 AB, Koobi Fora, Kenya, Using High-Resolution Spatial Analysis and FTIR Spectrometry|journal=Current Anthropology|volume=58|pages=S243–S257|doi=10.1086/692530|s2cid=148948219}}</ref><ref name=Gowlett2016/><ref name=Roebroeks2011/> and GnJi 1/6E in the [[Chemoigut Formation]], as far back as 1.5 Mya,<ref name=Gowlett2016/><ref name=Roebroeks2011/> and in South Africa, [[Wonderwerk Cave]], 1.7 Mya.<ref name=Beaumont2011>{{cite journal | vauthors = Beaumont PB |year=2011|title=The Edge: More on Fire-Making by about 1.7 Million Years Ago at Wonderwerk Cave in South Africa|journal=Current Anthropology|volume=52|issue=4|pages=585–595|doi=10.1086/660919|s2cid=144176681}}</ref> The first firekeepers are thought to have simply transported to caves and maintained naturally occurring fires for extended periods of time or only sporadically when the opportunity arose. Maintaining fires would require firekeepers to have knowledge of slow-burning materials such as dung.<ref name=Gowlett2016>{{cite journal | vauthors = Gowlett JA | title = The discovery of fire by humans: a long and convoluted process | journal = Philosophical Transactions of the Royal Society of London. Series B, Biological Sciences | volume = 371 | issue = 1696 | pages = 20150164 | date = June 2016 | pmid = 27216521 | pmc = 4874402 | doi = 10.1098/rstb.2015.0164 }}</ref> Fire becomes markedly more abundant in the wider archaeological record after 400,000–300,000 years ago, which can be explained as some advancement in fire management techniques took place at this time<ref name=Gowlett2016/> or human ancestors only opportunistically used fire until this time.<ref name=Roebroeks2011>{{cite journal | vauthors = Roebroeks W, Villa P | title = On the earliest evidence for habitual use of fire in Europe | journal = Proceedings of the National Academy of Sciences of the United States of America | volume = 108 | issue = 13 | pages = 5209–5214 | date = March 2011 | pmid = 21402905 | pmc = 3069174 | doi = 10.1073/pnas.1018116108 | doi-access = free | bibcode = 2011PNAS..108.5209R }}</ref><ref name=Sandgathe2017/><ref name=Ungar2006/><ref name=BenDor2011/> It is possible that firestarting was invented and lost and reinvented multiple times and independently by different communities rather than being invented in one place and spreading throughout the world.<ref name=Sandgathe2017>{{cite journal| vauthors = Sandgathe D |year=2017|title=Identifying and Describing Pattern and Process in the Evolution of Hominin Use of Fire|journal=Current Anthropology|volume=58|pages=S360–S370|doi=10.1086/691459|url=https://www.researchgate.net/publication/317042963|hdl=11858/00-001M-0000-002C-0141-3|s2cid=165025762|hdl-access=free}}</ref> The earliest evidence of hearths comes from Gesher Benot Ya'aqov, Israel, over 700,000 years ago, where fire is recorded in multiple layers in an area close to water, both uncharacteristic of natural fires.<ref name=Gowlett2013/> |
|||
Artificial lighting may have led to increased waking hours—modern humans have about a 16-hour waking period, whereas other apes are generally awake from only sunup to sundown—and these additional hours were probably used for socializing. Because of this, fire usage is probably also linked to the [[origin of language]].<ref name=Gowlett2016/><ref name=Gowlett2013/> Artificial lighting may have also made sleeping on the ground instead of the trees possible by keeping terrestrial predators at bay.<ref name=Gowlett2013/> |
|||
Migration into the frigid climate of Ice Age Europe may have only been possible because of fire, but evidence of fire usage in Europe until about 400–300,000 years ago is notably absent.<ref name=Roebroeks2011/> If these early European ''H. erectus'' did not have fire, it is largely unclear how they stayed warm, avoided predators, and prepared animal fat and meat for consumption. There was also a lower likelihood of naturally occurring fires due to lightning being less common in areas further north. It is possible that they only knew how to maintain fires in certain settings in the landscapes and prepared food some distance away from home, meaning evidence of fire and evidence of hominin activity are spaced far apart.<ref name=Gowlett2013>{{cite journal| vauthors = Gowlett JA, Wrangham RW |year=2013|title=Earliest fire in Africa: Towards the convergence of archaeological evidence and the cooking hypothesis|journal=Azania: Archaeological Research in Africa|volume=48|issue=1|pages=5–30|doi=10.1080/0067270X.2012.756754|s2cid=163033909|url=https://www.researchgate.net/publication/271530765}}</ref> Alternatively, ''H. erectus'' may have only pushed farther north during warmer [[interglacial]] periods—thus not requiring fire, food storage, or clothing technology—<ref>{{cite journal | vauthors = Antón SC | title = Natural history of Homo erectus | journal = American Journal of Physical Anthropology | volume = 122 | issue = S37 | pages = 126–170 | year = 2003 | pmid = 14666536 | doi = 10.1002/ajpa.10399 | doi-access = free }}</ref> and their dispersal patterns indicate they generally stayed in warmer lower-to-middle latitudes.<ref name=Carotenuto2016>{{cite journal | vauthors = Carotenuto F, Tsikaridze N, Rook L, Lordkipanidze D, Longo L, Condemi S, Raia P | title = Venturing out safely: The biogeography of Homo erectus dispersal out of Africa | journal = Journal of Human Evolution | volume = 95 | pages = 1–12 | date = June 2016 | pmid = 27260171 | doi = 10.1016/j.jhevol.2016.02.005 | bibcode = 2016JHumE..95....1C | hdl = 10356/82274 | hdl-access = free }}</ref> It is debated if the ''H. e. pekinensis'' inhabitants of [[Zhoukoudian]], Northern China, were capable of controlling fires as early as 770 kya to stay warm in what may have been a relatively cold climate.<ref>{{cite journal| vauthors = Zhong M, Shi C, Gao X, Wu X, Chen F, Zhang S, Zhang X, Olsen JW |year=2013|title=On the possible use of fire by ''Homo erectus'' at Zhoukoudian, China|journal=Chinese Science Bulletin|volume=59|issue=3|pages=335–343|doi=10.1007/s11434-013-0061-0|s2cid=93590269|url=https://www.researchgate.net/publication/272016716}}</ref> |
|||
==== Construction ==== |
|||
[[File:Terra-Amata-Hut.gif|thumb|Reconstruction of a [[Terra Amata (archaeological site)|Terra Amata]] dwelling<ref>{{cite web|url=https://www.musee-terra-amata.org/musee/le-site-acheuleen-de-terra-amata/ |title=Le site acheuléen de Terra Amata |language=fr |trans-title=The Acheulean site of Terra Amata |author=Musée de Préhistoire Terra Amata |website=Musée de Préhistoire Terra Amata |access-date=10 June 2022}}</ref>]] |
|||
In 1962, a {{cvt|12x14x1|ft|cm|order=flip}} circle made with volcanic rocks was discovered in [[Olduvai Gorge]]. At {{cvt|2–2.5|ft|cm|order=flip|adj=on}} intervals, rocks were piled up to {{cvt|6–9|in|cm|order=flip}} high. British palaeoanthropologist [[Mary Leakey]] suggested the rock piles were used to support poles stuck into the ground, possibly to support a [[windbreak]] or a rough hut. Some modern-day nomadic tribes build similar low-lying rock walls to build temporary shelters upon, bending upright branches as poles and using grasses or animal hide as a screen.<ref>{{cite book|url={{google books|plainurl=yes|id=eepULHufmF8C|page=24}} |vauthors=Leakey MD |author-link=Mary Leakey |year=1971 |title=Olduvai Gorge: Volume 3, Excavations in Beds I and II, 1960-1963 |publisher=[[Cambridge University Press]] |page=24 |isbn=9780521077231}}</ref> Dating to 1.75 Mya, it is the oldest claimed evidence of architecture.<ref>{{cite book|vauthors=Ingold T |year=2000 |title=The Perception of the Environment: Essays on Livelihood, Dwelling and Skill |chapter=Building, dwelling, living: how animals and people make themselves at home in the world |publisher=Psychology Press |page=184 |url={{google books|plainurl=yes|id=S3GakE5OT-kC|page=184}} |isbn=9780415228329}}</ref> |
|||
The earliest evidence of cave habitation is Wonderwerk Cave, South Africa, about 1.6 Mya, but evidence of cave use globally is sporadic until about 600 kya.<ref>{{cite journal|url=https://www.researchgate.net/publication/303107882 |vauthors=Ullman M, Hovers E, Goren-Inbar N, Frumkin A |year=2013 |title=Levantine cave dwellers: geographic and environmental aspects of early humans use of caves, case study from Wadi Amud, northern Israel |journal=International Congress of Speleology |volume=1}}</ref> |
|||
====Clothing==== |
|||
[[File:Homo-erectus Turkana-Boy (Ausschnitt) Fundort Nariokotome, Kenia, Rekonstruktion im Neanderthal Museum.jpg|thumb|upright=1.3|left|Reconstruction of Turkana boy with light clothing by Adrie and Alfons Kennis at the [[Neanderthal Museum]]]] |
|||
It is largely unclear when clothing was invented, with the earliest estimate stretching as far back as 3 Mya to compensate for a lack of insulating body hair.<ref name=Gilligan2010>{{cite journal| vauthors = Gilligan I |year=2010|title=The Prehistoric Development of Clothing: Archaeological Implications of a Thermal Model|journal=Journal of Archaeological Method and Theory |volume=15|pages=15–80|doi=10.1007/s10816-009-9076-x|s2cid=143004288}}</ref> It is known that [[head lice]] and [[body lice]] (the latter can only inhabit clothed individuals) for modern humans diverged about 170 kya, well before modern humans left Africa, meaning clothes were already well in use before encountering cold climates. One of the first uses of animal hide is thought to have been for clothing, and the oldest hide scrapers date to about 780 kya; however, this is not indicative of clothing.<ref>{{cite journal | vauthors = Toups MA, Kitchen A, Light JE, Reed DL | title = Origin of clothing lice indicates early clothing use by anatomically modern humans in Africa | journal = Molecular Biology and Evolution | volume = 28 | issue = 1 | pages = 29–32 | date = January 2011 | pmid = 20823373 | pmc = 3002236 | doi = 10.1093/molbev/msq234 }}</ref> |
|||
====Possible seafaring==== |
|||
Acheulean artifacts discovered on isolated islands that were never connected to land in the Pleistocene may show seafaring by ''H. erectus'' as early as 1 Mya in Indonesia. They had arrived on the islands of [[Flores]], [[Timor]], and [[Rote Island|Roti]], which would have necessitated crossing the [[Lombok Strait]] (the [[Wallace Line]]), at least before 800 kya. It is also possible they were the first European mariners as well and crossed the [[Strait of Gibraltar]] between North Africa and Spain. A 2021 genetic analysis of these island populations of ''H. erectus'' found no evidence of interbreeding with modern humans.<ref>{{Cite web|date=23 March 2021|title=New evidence in search for the mysterious Denisovans|url=https://www.sciencedaily.com/releases/2021/03/210323084732.htm|access-date=30 March 2021|website=ScienceDaily|language=en}}</ref> Seafaring capability would show ''H. erectus'' had a great capacity for planning, likely months in advance of the trip.<ref name=Bednarik1999>{{cite journal| vauthors = Bednarik RG |year=1999 |title= Pleistocene seafaring in the Mediterranean|journal=Anthropologie|volume=37|issue=3|pages=275–282|jstor=26294895}}</ref><ref>{{cite journal| vauthors = Bednarik RG |year=1998|title=An experiment in Pleistocene seafaring|journal=The International Journal of Nautical Archaeology|volume=27|issue=2|pages=139–149|doi=10.1111/j.1095-9270.1998.tb00797.x|bibcode=1998IJNAr..27..139B |url=http://www.ifrao.com/wp-content/uploads/2015/03/98Nautuical.pdf}}</ref> However, there is no evidence of the watercrafts required for such seafaring, though these would likely not survive in the fossil record. Additionally, experiments by Bednarik that show modern humans can make complex boats using materials available to ''H. erectus,'' actually undercut this hypothesis. Such experiments presuppose cognition rivalling ''H. sapiens'', despite the lack of evidence for such a premise. Instead, it may well be that ''Homo erectus'' reached Flores through [[Oceanic dispersal|natural rafts]] of vegetation that had been swept out to sea following a cyclone or tsunami.<ref name=":6" /> |
|||
Similarly, ''[[Homo luzonensis]]'' is dated between 771,000 and 631,000 years ago. Because Luzon has always been an island in the Quaternary, the ancestors of ''H. luzonensis'' would have had to have made a substantial sea crossing and crossed the [[Huxley Line]].<ref name="Détroit2019">{{cite journal | vauthors = Détroit F, Mijares AS, Corny J, Daver G, Zanolli C, Dizon E, Robles E, Grün R, Piper PJ | display-authors = 6 | title = A new species of Homo from the Late Pleistocene of the Philippines | journal = Nature | volume = 568 | issue = 7751 | pages = 181–186 | date = April 2019 | pmid = 30971845 | doi = 10.1038/s41586-019-1067-9 | name-list-style = amp | s2cid = 106411053 | bibcode = 2019Natur.568..181D | url = https://hal.archives-ouvertes.fr/hal-02296712/file/Detroit_%26_al_2019_Nature_postprint.pdf }}</ref> |
|||
====Healthcare==== |
|||
[[File:Dmanisi fossils D 3444 + D 3900 (Replika).jpg|thumb|upright|Skull of a toothless ''H. e. georgicus'']] |
|||
The earliest probable example of infirming sick group members is a 1.77 Mya ''H. e. georgicus'' specimen who had lost all but one tooth due to age or [[gum disease]], the earliest example of severe chewing impairment, yet still survived for several years afterwards. However, it is possible australopithecines were capable of caring for debilitated group members.<ref>{{cite journal| vauthors = Spikins P, Needham A, Wright B, Dytham C, Gatta M, Hitchens G |year=2019 |title= Living to fight another day: The ecological and evolutionary significance of Neanderthal healthcare|journal=Quaternary Science Reviews|volume=217|pages=98–118|doi=10.1016/j.quascirev.2018.08.011|bibcode=2019QSRv..217...98S|doi-access=free}}</ref> Unable to chew, this ''H. e. georgicus'' individual probably ate soft plant or animal foods possibly with assistance from other group members. High-latitude groups are thought to have been predominantly carnivorous, eating soft tissue such as [[bone marrow]] or brains, which may have increased survival rates for toothless individuals.<ref>{{cite journal | vauthors = Lordkipanidze D, Vekua A, Ferring R, Rightmire GP, Agusti J, Kiladze G, Mouskhelishvili A, Nioradze M, Ponce de León MS, Tappen M, Zollikofer CP | display-authors = 6 | title = Anthropology: the earliest toothless hominin skull | journal = Nature | volume = 434 | issue = 7034 | pages = 717–718 | date = April 2005 | pmid = 15815618 | doi = 10.1038/434717b | bibcode = 2005Natur.434..717L | s2cid = 52800194 }}</ref> |
|||
The 1.5 Mya Turkana boy was diagnosed with juvenile [[spinal disc herniation]], and, because this specimen was still growing, this caused some [[scoliosis]] (abnormal curving of the spine). These usually cause recurrent lower back pain and [[sciatica]] (pain running down the leg), and likely restricted Turkana boy in walking, bending, and other daily activities. The specimen appears to have survived into adolescence, which evidences advanced group care.<ref>{{cite journal | vauthors = Haeusler M, Schiess R, Boeni T | title = Evidence for juvenile disc herniation in a homo erectus boy skeleton | journal = Spine | volume = 38 | issue = 3 | pages = E123–E128 | date = February 2013 | pmid = 23154836 | doi = 10.1097/BRS.0b013e31827cd245 | s2cid = 11534863 | url = https://www.zora.uzh.ch/id/eprint/76396/1/Haeusler_et_al_Evidence_for_juvenile_disc_herniation.pdf }}</ref> |
|||
The 1,000–700 kya Java man specimen presents a noticeable [[osteocyte]] on the femur, likely [[Paget's disease of bone]], and [[osteopetrosis]], thickening of the bone, likely resulting from [[skeletal fluorosis]] caused by ingestion of food contaminated by fluorine-filled volcanic ash (as the specimen was found in ash-filled [[stratum (geology)|strata]]). Livestock that grazes on volcanic ash ridden fields typically die of acute intoxication within a few days or weeks.<ref>{{cite journal | vauthors = Soriano M | title = The fluoric origin of the bone lesion in the Pithecanthropus erectus femur | journal = American Journal of Physical Anthropology | volume = 32 | issue = 1 | pages = 49–57 | date = January 1970 | pmid = 4984453 | doi = 10.1002/ajpa.1330320107 }}</ref> |
|||
=== Art and rituals === |
|||
{{See also|Prehistoric art}} |
|||
{{Multiple image|width=300|direction=vertical|image1=Homo Erectus shell with geometric incisions circa 500,000 BP, Naturalis Biodiversity Center, Netherlands (with detail).jpg|caption1=Engraved [[Pseudodon shell DUB1006-fL]] from [[Trinil]], [[Java]]|image2=Museo de la Evolucion Humana Burgos - Tan Tan and Berekhat Ram Pebbles.jpg|caption2=Replicas of the "[[Venus of Tan-Tan]]" (left) and "[[Venus of Berekhat Ram]]" (right)}} |
|||
An engraved [[Pseudodon shell DUB1006-fL]] with geometric markings could possibly be evidence of the earliest art-making, dating back to 546–436 kya. Art-making capabilities could be considered evidence of symbolic thinking, which is associated with modern cognition and behavior.<ref name=":1">{{cite journal | vauthors = Joordens JC, d'Errico F, Wesselingh FP, Munro S, de Vos J, Wallinga J, Ankjærgaard C, Reimann T, Wijbrans JR, Kuiper KF, Mücher HJ, Coqueugniot H, Prié V, Joosten I, van Os B, Schulp AS, Panuel M, van der Haas V, Lustenhouwer W, Reijmer JJ, Roebroeks W | display-authors = 6 | title = Homo erectus at Trinil on Java used shells for tool production and engraving | journal = Nature | volume = 518 | issue = 7538 | pages = 228–231 | date = February 2015 | pmid = 25470048 | doi = 10.1038/nature13962 | s2cid = 4461751 | bibcode = 2015Natur.518..228J }}</ref><ref>{{cite journal | vauthors = Henshilwood CS, d'Errico F, Watts I | title = Engraved ochres from the Middle Stone Age levels at Blombos Cave, South Africa | journal = Journal of Human Evolution | volume = 57 | issue = 1 | pages = 27–47 | date = July 2009 | pmid = 19487016 | doi = 10.1016/j.jhevol.2009.01.005 | bibcode = 2009JHumE..57...27H }}</ref><ref>{{cite journal | vauthors = d'Errico F, Moreno RG, Rifkin RF | year = 2012 | title = Technological, elemental and colorimetric analysis of an engraved ochre fragment from the Middle Stone Age levels of Klasies River Cave 1, South Africa | journal = J. Archaeol. Sci. | volume = 39 | issue = 4| pages = 942–952 | doi = 10.1016/j.jas.2011.10.032 | bibcode = 2012JArSc..39..942D }}</ref><ref name="Nature Article">{{cite journal | vauthors = Callaway E |title=''Homo erectus'' made world's oldest doodle 500,000 years ago |journal=Nature News |doi=10.1038/nature.2014.16477 |url=https://www.nature.com/news/homo-erectus-made-world-s-oldest-doodle-500-000-years-ago-1.16477|year=2014 |s2cid=164153158 }}</ref> In 1976, American archeologist [[Alexander Marshack]] asserted that engraved lines on an ox rib, associated with Acheulean lithics, from [[Pech de l'Azé]], France, are similar to a [[meander (art)|meander design]] found in modern human Upper Paleolithic cave art.<ref name=Dickson1992>{{cite book| vauthors = Dickson DB |year=1992|title=The Dawn of Belief: Religion in the Upper Paleolithic of Southwestern Europe|publisher=University of Arizona Press|pages=40–46|url={{google books|plainurl=yes|id=DNr5YIygjMMC|page=40}}|isbn=978-0-8165-1336-9}}</ref> Three [[ostrich eggshell beads]] associated with Achuelian lithics were found in northwestern Africa, the earliest disc beads ever found, and Acheulian disc beads have also been found in France and Israel.<ref name=Bednarik1999/> The Middle Pleistocene "[[Venus of Tan-Tan]]" and "[[Venus of Berekhat Ram]]" are postulated to been crafted by ''H. erectus'' to resemble a human form. They were mostly formed by natural weathering, but slightly modified to emphasize certain grooves to suggest hairline, limbs, and eyes.<ref name=Morriss2009/><ref>{{cite journal| vauthors = d'Errico F, Nowell A |year=2000|title=A New Look at the Berekhat Ram Figurine: Implications for the Origins of Symbolism|journal=Cambridge Archaeological Journal|volume=10|issue=1|pages=123–167|doi=10.1017/S0959774300000056|s2cid=163138037}}</ref> The former has traces of pigments on the front side, possibly indicating it was colored.<ref name=Morriss2009>{{cite journal | vauthors = Morriss-Kay GM | title = The evolution of human artistic creativity | journal = Journal of Anatomy | volume = 216 | issue = 2 | pages = 158–176 | date = February 2010 | pmid = 19900185 | pmc = 2815939 | doi = 10.1111/j.1469-7580.2009.01160.x }}</ref> |
|||
''H. erectus'' was also the earliest human to have intentionally collected red-colored pigments, namely [[ochre]], recorded as early as the Middle Pleistocene. Ochre lumps at [[Olduvai Gorge]], Tanzania—associated with the 1.4 Ma [[Olduvai Hominid 9]]—and [[Ambrona]], Spain—which dates to 424–374 kya—were suggested to have been struck by a hammerstone and purposefully shaped and trimmed.<ref name=Watts2014/><ref name=Dickson1992/> At Terra Amata, France—which dates to 425–400 or 355–325 kya—red, yellow, and brown ochres were recovered in association with pole structures; ochre was probably heated to achieve such a wide color range.<ref name=Watts2014>{{cite book| vauthors = Watts I |year=2014|chapter=The red thread: pigment use and the evolution of collective ritual|title=The Social Origins of Language|publisher=Oxford University Press|pages=222–223|url={{google books|plainurl=yes|id=0HtYCwAAQBAJ|page=222}}|isbn=978-0-19-966533-4}}</ref><ref>{{cite book| vauthors = de Lumley H, Boone Y |year=1976|chapter=Les structures d'habitat au Paléolithique moyen|trans-chapter=Housing structures from the lower Paleolithic|title=La Préhistoire française: Les civilisations paléolithiques et mésolithiques de la France|trans-title=French prehistory: the Paleolithic and Mesolithic civilizations of France| veditors = de Lumley H, Guilaine J |publisher=Éditions du Centre national de la recherche scientifique|isbn=978-2-222-01968-8}}</ref> As it is unclear if ''H. erectus'' could have used ochre for any practical application, ochre collection might indicate that ''H. erectus'' was the earliest human to have exhibited a sense of [[aesthetics]] and to think beyond simply survival. Later human species are postulated to have used ochre as body paint, but in the case of ''H. erectus'', it is contested if body paint was used so early in time. Further, it is unclear if these few examples are not simply isolated incidents of ochre use, as ochre is much more prevalent in Middle and Upper Paleolithic sites attributed to Neanderthals and ''H. sapiens''.<ref name=Wreschner1980>{{cite journal|url=http://sites.utexas.edu/butzer/files/2017/03/Wreschner_Butzer-1980-RedOchre.pdf| vauthors = Wreschner EE, Bolton R, Butzer KW, Delporte H, Häusler A, Heinrich A, Jacobson-Widding A, Malinowski T, Masset C, Miller SF, Ronen A | display-authors = 6 |year=1980|title=Red Ochre and Human Evolution: A Case for Discussion|journal=Current Anthropology|volume=21|issue=5|pages=632–633<!--only citing these pages-->|doi=10.1086/202541|jstor=2741829|s2cid=88099778}}</ref><ref name=Dickson1992/> |
|||
In 1935, Jewish-German anthropologist [[Franz Weidenreich]] speculated that the inhabitants of the Chinese [[Zhoukoudian|Zhoukoudian Peking Man site]] were members of some Lower Paleolithic Skull Cult because the skulls all showed fatal blows to the head, breaking in of the [[foramen magnum]] at the base of the skull, by-and-large lack of preserved facial aspects, an apparently consistent pattern of breaking on the mandible, and a lack of post-cranial remains (elements that are not the skull). He believed that the inhabitants were [[headhunting|headhunters]], and smashed open the skulls and ate the brains of their victims.<ref>{{cite journal| vauthors = Weidenreich F |author-link=Franz Weidenreich|year=1935|title=The ''Sinanthropus'' Population of Choukoutien (Locality 1) with a Preliminary Report on New Discoveries|journal=Bulletin of the Geological Society of China|volume=14|issue=4|pages=427–468|doi=10.1111/j.1755-6724.1935.mp14004001.x}}</ref><ref name=Dickson1992/> However, scavenging animals and natural forces such as flooding can also inflict the same kind of damage to skulls,<ref name=Dickson1992/> and there is not enough evidence to suggest manhunting or cannibalism.<ref>{{cite journal| vauthors = Binford LR, Ho CK |author-link=Lewis Binford |year=1985|title=Taphonomy at a Distance: Zhoukoudian, 'The Cave Home of Beijing Man'?|journal=Current Anthropology|volume=26|issue=4|pages=413–442|doi=10.1086/203303|jstor=2742759|s2cid=147164100}}</ref> |
|||
In 1999, British science writers [[Marek Kohn]] and [[Steven Mithen]] said that many hand axes exhibit no wear and were produced en masse, and concluded that these symmetrical, tear-drop shaped lithics functioned primarily as [[display (zoology)|display]] tools so males could prove their fitness to females in some courting ritual, and were discarded afterwards.<ref>{{cite journal| vauthors = Kohn M, Mithen S |author-link=Marek Kohn|author2-link=Steven Mithen|year=1999|title=Handaxes: products of sexual selection?|journal=Antiquity|volume=73|issue=281|pages=518–526|doi=10.1017/S0003598X00065078|s2cid=162903453}}</ref> However, an apparent lack of reported wearing is likely due to a lack of use-wear studies, and only a few sites yield an exorbitant sum of hand axes likely due to gradual accumulation over generations instead of mass production.<ref name=Nowell2009>{{cite journal| vauthors = Nowell A, Chang ML |year=2009|title=The Case Against Sexual Selection as an Explanation of Handaxe Morphology |journal=PaleoAnthropology |pages=77–88|url=http://paleoanthro.reedd.webfactional.com/enwiki/static/journal/content/PA20090077.pdf}}</ref> |
|||
===Language=== |
|||
In 1984, the vertebral column of the 1.6 Mya adolescent [[Turkana boy]] indicated that this individual did not have properly developed respiratory muscles in order to produce speech. In 2001, American anthropologists Bruce Latimer and James Ohman concluded that Turkana boy was afflicted by [[skeletal dysplasia]] and [[scoliosis]].<ref>{{cite journal| vauthors = Latimer B, Ohman J |year=2001|title=Axial dysplasia in ''Homo erectus''|journal=Journal of Human Evolution|volume=40}}</ref> In 2006, American anthropologist [[Marc Meyer]] and colleagues described a 1.8 Mya ''H. e. georgicus'' specimen as having a spine within the range of variation of modern human spines, contending that Turkana boy had [[spinal stenosis]] and was thus not representative of the species. Also, because he considered ''H. e. georgicus'' ancestral to all non-African ''H. erectus'', Meyer concluded that the respiratory muscles of all ''H. erectus'' (at least non-''H. ergaster'') would not have impeded vocalisation or speech production.<ref>{{cite conference | vauthors = Meyer M, Lordkipanidze D, Vekua A | title = Language and empathy in Homo erectus: Behaviors suggested by a modern spinal cord from Dmanisi, but not Nariokotome. | conference = Annual meeting of the Paleoanthroplogy Society | location = San Juan, Puerto Rico | date = 2006 | url = https://www.researchgate.net/publication/263198625 }}</ref> However, in 2013 and 2014, anthropologist Regula Schiess and colleagues concluded that there is no evidence of any congenital defects in Turkana boy, and considered the specimen representative of the species.<ref>{{cite journal | vauthors = Schiess R, Haeusler M | title = No skeletal dysplasia in the Nariokotome boy KNM-WT 15000 (Homo erectus)--a reassessment of congenital pathologies of the vertebral column | journal = American Journal of Physical Anthropology | volume = 150 | issue = 3 | pages = 365–374 | date = March 2013 | pmid = 23283736 | doi = 10.1002/ajpa.22211 }}</ref><ref>{{cite journal | vauthors = Schiess R, Boeni T, Rühli F, Haeusler M | title = Revisiting scoliosis in the KNM-WT 15000 Homo erectus skeleton | journal = Journal of Human Evolution | volume = 67 | issue = 48–59 | pages = 48–59 | date = February 2014 | pmid = 24491377 | doi = 10.1016/j.jhevol.2013.12.009 | bibcode = 2014JHumE..67...48S | url = https://www.zora.uzh.ch/id/eprint/92705/1/Schiess_et_al._-_Scoliosis_in_WT15000_2013-10-24.pdf }}</ref> |
|||
Neurologically, all ''Homo'' have similarly configured brains, and, likewise, the [[Broca's area|Broca's]] and [[Wernicke's area|Wernicke's]] areas (in charge of sentence formulation and speech production in modern humans) of ''H. erectus'' were comparable to those of modern humans. However, this is not indicative of anything in terms of speech capability as even large chimpanzees can have similarly expanded Broca's area, and it is unclear if these areas served as language centers in archaic humans.<ref>{{cite book| vauthors = Luef EM |year=2018|chapter=Tracing the human brain's classical language areas in extant and extinct hominids|title=The talking species: Perspectives on the evolutionary, neuronal and cultural foundations of language|publisher=Uni-Press Graz|url=https://www.researchgate.net/publication/327285824|isbn=978-3-902666-52-9}}</ref> A 1-year-old ''H. erectus'' specimen shows that an extended childhood to allow for brain growth, which is a prerequisite in language acquisition, was not exhibited in this species.<ref name=Coqueugniot2004/> |
|||
The [[hyoid bone]] supports the tongue and makes possible modulation of the [[vocal tract]] to control pitch and volume. A 400 kya ''H. erectus'' hyoid bone from [[Castel di Guido]], Italy, is bar-shaped—more similar to that of other ''Homo'' than to that of non-human apes and ''Australopithecus''—but is devoid of muscle impressions, has a shield-shaped body, and is implied to have had reduced greater horns, meaning ''H. erectus'' lacked a humanlike vocal apparatus and thus anatomical prerequisites for a modern human level of speech.<ref>{{cite journal | vauthors = Capasso L, Michetti E, D'Anastasio R | title = A Homo erectus hyoid bone: possible implications for the origin of the human capability for speech | journal = Collegium Antropologicum | volume = 32 | issue = 4 | pages = 1007–1011 | date = December 2008 | pmid = 19149203 }}</ref> Increasing brain size and cultural complexity in tandem with technological refinement, and the hypothesis that articulate Neanderthals and modern humans may have inherited speech capabilities from the last common ancestor, could possibly indicate that ''H. erectus'' used some [[Origin of language|proto-language]] and built the basic framework which fully fledged languages would eventually be built around.<ref>{{cite journal | vauthors = Hillert DG | title = On the Evolving Biology of Language | journal = Frontiers in Psychology | volume = 6 | pages = 1796 | year = 2015 | pmid = 26635694 | pmc = 4656830 | doi = 10.3389/fpsyg.2015.01796 | doi-access = free }}</ref> However, this ancestor may have instead been ''H. heidelbergensis'', as a hyoid bone of a 530 kya ''H. heidelbergensis'' specimen from the Spanish [[Sima de los Huesos]] Cave is like that of modern humans,<ref>{{cite journal | vauthors = Martínez I, Arsuaga JL, Quam R, Carretero JM, Gracia A, Rodríguez L | title = Human hyoid bones from the middle Pleistocene site of the Sima de los Huesos (Sierra de Atapuerca, Spain) | journal = Journal of Human Evolution | volume = 54 | issue = 1 | pages = 118–124 | date = January 2008 | pmid = 17804038 | doi = 10.1016/j.jhevol.2007.07.006 | bibcode = 2008JHumE..54..118M | url = https://eprints.ucm.es/26853/1/1-s2.0-S0047_1.pdf }}</ref> and another specimen from the same area shows an auditory capacity sensitive enough to pick up human speech.<ref>{{cite journal | vauthors = Martínez I, Rosa M, Arsuaga JL, Jarabo P, Quam R, Lorenzo C, Gracia A, Carretero JM, Bermúdez de Castro JM, Carbonell E | display-authors = 6 | title = Auditory capacities in Middle Pleistocene humans from the Sierra de Atapuerca in Spain | journal = Proceedings of the National Academy of Sciences of the United States of America | volume = 101 | issue = 27 | pages = 9976–9981 | date = July 2004 | pmid = 15213327 | pmc = 454200 | doi = 10.1073/pnas.0403595101 | doi-access = free | bibcode = 2004PNAS..101.9976M }}</ref> |
|||
==Extinction== |
|||
{{Expand section|date=December 2020}} |
|||
The last known occurrence of ''Homo erectus'' is 117,000–108,000 years ago in [[Ngandong]], [[Java (island)|Java]] according to a study published in 2019.<ref name=Rizal/> |
|||
In 2020, researchers reported that ''Homo erectus'' and ''[[Homo heidelbergensis]]'' lost more than half of their climate [[Ecological niche|niche]] – climate they were adapted to – with no corresponding reduction in physical range, just before [[extinction]] and that climate change played a substantial role in extinctions of past ''Homo'' species.<ref>{{cite news | vauthors = Padmanaban D |title=Climate Change May Have Been a Major Driver of Ancient Hominin Extinctions |url=https://www.sapiens.org/archaeology/hominin-extinctions/ |website=SAPIENS |access-date=9 November 2020 |date=6 November 2020}}</ref><ref>{{cite news |title=Climate change likely drove early human species to extinction, modeling study suggests |url=https://phys.org/news/2020-10-climate-drove-early-human-species.html |access-date=9 November 2020 |work=phys.org |language=en}}</ref><ref>{{cite journal | vauthors = Raia P, Mondanaro A, Melchionna M, Di Febbraro M, Diniz-Filho JA, Rangel TF, Holden PB, Carotenuto F, Edwards NR, Lima-Ribeiro MS, Profico A, Maiorano L, Castiglione S, Serio C, Rook L | display-authors = 6 |title=Past Extinctions of Homo Species Coincided with Increased Vulnerability to Climatic Change |journal=One Earth |date=23 October 2020 |volume=3 |issue=4 |pages=480–490 |doi=10.1016/j.oneear.2020.09.007 | bibcode = 2020OEart...3..480R |hdl=2158/1211341 |language=en |issn=2590-3330|doi-access=free |hdl-access=free }}</ref> |
|||
==Fossils== |
|||
[[File:Homo erectus KNM ER 3733.jpg|thumb|Homo erectus KNM ER 3733 actual skull]] |
|||
The lower cave of China's Zhoukoudian Cave is one of the most important archaeological sites worldwide.<ref name=":2">Zanolli, Clément, et al. "Inner Tooth Morphology of Homo Erectus from Zhoukoudian. New Evidence from an Old Collection Housed at Uppsala University, Sweden." ''Journal of Human Evolution'', vol. 116, Mar. 2018, pp. 1–13.</ref> There have been remains of 45 ''Homo erectus'' individuals found and thousands of tools recovered.<ref name=":2" /> Most of these remains were lost during World War 2, with the exception of two postcranial elements that were rediscovered in China in 1951 and four human teeth from 'Dragon Bone Hill'.<ref name=":2" /> |
|||
New evidence has shown that ''Homo erectus'' does not have uniquely thick vault bones, as was previously thought.<ref name=":3">Copes, Lynn E., and William H. Kimbel. "Cranial Vault Thickness in Primates: Homo Erectus Does Not Have Uniquely Thick Vault Bones." ''Journal of Human Evolution'', vol. 90, Jan. 2016, pp. 120–134.</ref> Testing showed that neither Asian nor African ''Homo erectus'' had uniquely large vault bones.<ref name=":3" /> |
|||
=== Individual fossils === |
|||
Some of the major ''Homo erectus'' fossils: |
Some of the major ''Homo erectus'' fossils: |
||
* Indonesia (island of Java): [[Trinil 2]] ([[holotype]]), [[Sangiran]] collection, Sambungmachan collection,<ref>{{cite journal |
* Indonesia (island of Java): [[Trinil 2]] ([[holotype]]), [[Sangiran]] collection, Sambungmachan collection,<ref>{{cite journal | vauthors = Delson E, Harvati K, Reddy D, Marcus LF, Mowbray K, Sawyer GJ, Jacob T, Márquez S | display-authors = 6 | title = The Sambungmacan 3 Homo erectus calvaria: a comparative morphometric and morphological analysis | journal = The Anatomical Record | volume = 262 | issue = 4 | pages = 380–397 | date = April 2001 | pmid = 11275970 | doi = 10.1002/ar.1048 | s2cid = 25438682 | doi-access = free }}</ref> [[Solo Man|Ngandong collection]] |
||
* China ("[[Peking Man]]"): Lantian (Gongwangling and Chenjiawo), Yunxian, [[Zhoukoudian]], Nanjing, [[Hexian]] |
* China ("[[Peking Man]]"): Lantian (Gongwangling and Chenjiawo), [[Xuetangliangzi|Yunxian]], [[Zhoukoudian]], Nanjing, [[Hexian]] |
||
* Kenya: [[KNM ER 3883]], [[KNM ER 3733]] |
* Kenya: [[KNM ER 3883]], [[KNM ER 3733]] |
||
* Vietnam: Northern, [[Tham Khuyen]],<ref>{{cite journal | vauthors = Ciochon R, Long VT, Larick R, González L, Grün R, de Vos J, Yonge C, Taylor L, Yoshida H, Reagan M | display-authors = 6 | title = Dated co-occurrence of Homo erectus and Gigantopithecus from Tham Khuyen Cave, Vietnam | journal = Proceedings of the National Academy of Sciences of the United States of America | volume = 93 | issue = 7 | pages = 3016–3020 | date = April 1996 | pmid = 8610161 | pmc = 39753 | doi = 10.1073/pnas.93.7.3016 | doi-access = free | bibcode = 1996PNAS...93.3016C }}</ref> Hoa Binh{{citation needed|date=October 2013}} |
|||
* Vértesszőlős, Hungary "[[Samu (Homo erectus)|Samu]]" |
|||
* Republic of Georgia: Dmanisi collection ("''[[Homo erectus georgicus]]''") |
|||
* Vietnam: Northern, Tham Khuyen,<ref>{{cite journal |vauthors=Ciochon R, Long VT, Larick R, etal |title=Dated co-occurrence of Homo erectus and Gigantopithecus from Tham Khuyen Cave, Vietnam |journal=Proceedings of the National Academy of Sciences of the United States of America |volume=93 |issue=7 |pages=3016–20 |date=April 1996 |pmid=8610161 |pmc=39753 |url=http://www.pnas.org/cgi/pmidlookup?view=long&pmid=8610161 |doi=10.1073/pnas.93.7.3016}}</ref> Hoa Binh{{citation needed|date=October 2013}} |
|||
* Republic of Georgia: Dmanisi collection ("[[Homo erectus georgicus]]") |
|||
* Ethiopia: [[Daka skull|Daka calvaria]] |
* Ethiopia: [[Daka skull|Daka calvaria]] |
||
* Eritrea: Buia cranium (possibly H. ergaster)<ref>http://archive.archaeology.org/9809/newsbriefs/eritrea.html |
* Eritrea: Buia cranium (possibly ''H. ergaster'')<ref>{{cite web|url= http://archive.archaeology.org/9809/newsbriefs/eritrea.html| title= New Skull from Eritrea| volume= 51 |number= 5| date= September–October 1998 | vauthors = Schuster AM | work= Archaeology| access-date= 3 October 2015}}</ref> |
||
* [[Denizli Province]], Turkey: Kocabas fossil<ref |
* [[Denizli Province]], Turkey: Kocabas fossil<ref name="First Homo erectus from Turkey and"/> |
||
* [[Drimolen]], South Africa: DNH 134<ref>{{cite journal | vauthors = Herries AI, Martin JM, Leece AB, Adams JW, Boschian G, Joannes-Boyau R, Edwards TR, Mallett T, Massey J, Murszewski A, Neubauer S, Pickering R, Strait DS, Armstrong BJ, Baker S, Caruana MV, Denham T, Hellstrom J, Moggi-Cecchi J, Mokobane S, Penzo-Kajewski P, Rovinsky DS, Schwartz GT, Stammers RC, Wilson C, Woodhead J, Menter C | display-authors = 6 | title = Contemporaneity of ''Australopithecus'', ''Paranthropus'', and early ''Homo erectus'' in South Africa | journal = Science | volume = 368 | issue = 6486 | pages = eaaw7293 | date = April 2020 | pmid = 32241925 | doi = 10.1126/science.aaw7293 | hdl-access = free | s2cid = 214763272 | hdl = 11568/1040368 }}</ref> |
|||
== |
==Gallery== |
||
<gallery mode="packed-hover" heights="120" class="center"> |
|||
{{Gallery |
|||
File:Homo erectus tautavelensis.jpg|''[[Homo erectus tautavelensis]]'' skull. |
|||
| lines = 3 |
|||
File:Tautavel UK 2.JPG|Replica of lower jaws of ''Homo erectus'' from [[Tautavel]], [[France]]. |
|||
| align = center |
|||
File:Calvaria Sangiran II (A).jpg|[[Calvaria (skull)|Calvaria]] "[[Sangiran]] II" original, collection [[Gustav Heinrich Ralph von Koenigswald|Koenigswald]], [[Senckenberg Museum]]. |
|||
|File:Homo erectus tautavelensis.jpg|''[[Homo erectus tautavelensis]]'' skull. |
|||
File:Homo erectus hand axe Daka Ethiopia.jpg|A reconstruction based on evidence from the [[Daka skull|Daka]] Member, Ethiopia |
|||
|File:Tautavel UK 2.JPG|Replica of lower jaws of ''Homo erectus'' from [[Tautavel]], [[France]]. |
|||
File:Pithecanthropus-erectus.jpg|Original fossils of ''Pithecanthropus erectus'' (now ''Homo erectus'') found in [[Java]] in 1891. |
|||
|File:Calvaria Sangiran II (A).jpg|[[Calvaria (skull)|Calvaria]] "[[Sangiran]] II" Original, Collection [[Gustav Heinrich Ralph von Koenigswald|Koenigswald]], [[Senckenberg Museum]]. |
|||
</gallery> |
|||
|File:Homo erectus hand axe Daka Ethiopia.jpg|A reconstruction based on evidence from the [[Daka skull|Daka]] Member, Ethiopia |
|||
}} |
|||
==See also== |
== See also == |
||
* ''[[ |
* ''[[Anthropopithecus]]'' |
||
* [[Kozarnika, Dimovo Municipality]] |
|||
* [[Java Man]] |
|||
* [[Multiregional origin of modern humans]] |
|||
* [[Kozarnika]] |
|||
'''General:''' |
'''General:''' |
||
* [[Dawn of Humanity (film)|''Dawn of Humanity'' (2015 PBS film)]] |
|||
* [[List of fossil sites]] ''(with link directory)'' |
* [[List of fossil sites]] ''(with link directory)'' |
||
* [[List of human evolution fossils]] ''(with images)'' |
* [[List of human evolution fossils]] ''(with images)'' |
||
==References== |
== References == |
||
{{Reflist |
{{Reflist}} |
||
== Further reading == |
|||
==External links== |
|||
* {{cite magazine|title=Homo Erectus Unearthed| vauthors = Leakey R, Walker A |magazine=[[National Geographic (magazine)|National Geographic]]|pages=624–629|volume=168|issue=5|date=November 1985|issn=0027-9358|oclc=643483454}} |
|||
*[http://www.bradshawfoundation.com/origins/homo_erectus.php Homo erectus] Origins - Exploring the Fossil Record - Bradshaw Foundation |
|||
* {{cite book| vauthors = Sigmon BA, Cybulski JS |year=1981|title=Homo erectus: Papers in Honor of Davidson Black|publisher=University of Toronto Press|jstor=10.3138/j.ctvcj2jdw.11|ref={{harvid|Sigmon|1981}}}} |
|||
{{Commons category|Homo erectus}} |
|||
* {{cite book| vauthors = Theunissen B, Theunissen LT |year=2012|title=Eugène Dubois and the Ape-Man from Java|publisher=Springer Netherlands|isbn=9789400922099|ref={{harvid|Theunissen|2012}}}} |
|||
*[http://www.archaeologyinfo.com/homoerectus.htm Archaeology Info] |
|||
*[http://humanorigins.si.edu/evidence/human-fossils/species/homo-erectus Homo erectus] – The Smithsonian Institution's Human Origins Program |
|||
*[http://news.bbc.co.uk/1/hi/sci/tech/6937476.stm Possible co-existence with Homo Habilis] – BBC News |
|||
*[[John D. Hawks|John Hawks]]'s [http://johnhawks.net/weblog/fossils/middle/kocabas/kappelman_2007_kocabas_tuberculosis.html discussion of the Kocabas fossil] |
|||
*[http://www-personal.une.edu.au/~pbrown3/palaeo.html Peter Brown's Australian and Asian Palaeoanthropology] |
|||
== External links == |
|||
{{Commons category|Homo erectus}}{{Wikispecies|Homo erectus|''Homo erectus''}} |
|||
* [http://www.bradshawfoundation.com/origins/homo_erectus.php Homo erectus] Origins – Exploring the Fossil Record – Bradshaw Foundation |
|||
* [http://www.archaeologyinfo.com/homoerectus.htm Archaeology Info] {{Webarchive|url=https://web.archive.org/web/20110516125629/http://www.archaeologyinfo.com/homoerectus.htm |date=16 May 2011 }} |
|||
* [http://humanorigins.si.edu/evidence/human-fossils/species/homo-erectus Homo erectus] – The Smithsonian Institution's Human Origins Program |
|||
* [http://news.bbc.co.uk/1/hi/sci/tech/6937476.stm Possible co-existence with Homo Habilis] – BBC News |
|||
* [[John D. Hawks|John Hawks]]'s [http://johnhawks.net/weblog/fossils/middle/kocabas/kappelman_2007_kocabas_tuberculosis.html discussion of the Kocabas fossil] |
|||
* [https://web.archive.org/web/20080511210522/http://www-personal.une.edu.au/~pbrown3/palaeo.html Peter Brown's Australian and Asian Palaeoanthropology] |
|||
* [http://atlasofhumanevolution.com/HomoErectus.asp The Age of Homo erectus] – Interactive Map of the Journey of Homo erectus out of Africa |
|||
* [http://humanorigins.si.edu/evidence/human-evolution-timeline-interactive Human Timeline (Interactive)] – [[Smithsonian Institution|Smithsonian]], [[National Museum of Natural History]] (August 2016). |
|||
{{Human Evolution}} |
{{Human Evolution}} |
||
{{Prehistoric technology}} |
{{Prehistoric technology}} |
||
{{Taxonbar|from=Q101362}} |
|||
{{Authority control}} |
{{Authority control}} |
||
{{portal bar|Evolutionary biology|Science}} |
|||
{{DEFAULTSORT:Homo Erectus}} |
{{DEFAULTSORT:Homo Erectus}} |
||
[[Category: |
[[Category:Homo erectus| ]] |
||
[[Category:Mammals described in 1892]] |
|||
[[Category:Pliocene primates]] |
[[Category:Pliocene primates]] |
||
[[Category:Pleistocene primates]] |
[[Category:Pleistocene primates]] |
||
[[Category:Pleistocene mammals of Africa]] |
[[Category:Pleistocene mammals of Africa]] |
||
[[Category:Fossil taxa described in 1892]] |
|||
[[Category:Prehistoric Indonesia]] |
[[Category:Prehistoric Indonesia]] |
||
[[Category:Prehistoric China]] |
[[Category:Prehistoric China]] |
||
| Line 280: | Line 322: | ||
[[Category:Prehistoric Anatolia]] |
[[Category:Prehistoric Anatolia]] |
||
[[Category:Prehistoric Spain]] |
[[Category:Prehistoric Spain]] |
||
[[Category: |
[[Category:Pleistocene mammals of Asia]] |
||
[[Category: |
[[Category:Pleistocene mammals of Europe]] |
||
[[Category:Taxa named by Eugène Dubois]] |
|||
[[Category:Tool-using mammals]] |
|||
[[Category:Early species of Homo]] |
|||
[[Category:Fossil taxa described in 1893]] |
|||
Latest revision as of 21:47, 19 December 2024
| Homo erectus | |
|---|---|
_presented_at_Paleozoological_Museum_of_China.jpg)
| |
| Replica of the skull of Peking Man at the Paleozoological Museum of China | |
| Scientific classification | |
| Domain: | Eukaryota |
| Kingdom: | Animalia |
| Phylum: | Chordata |
| Class: | Mammalia |
| Order: | Primates |
| Suborder: | Haplorhini |
| Infraorder: | Simiiformes |
| Family: | Hominidae |
| Subfamily: | Homininae |
| Tribe: | Hominini |
| Genus: | Homo |
| Species: | †H. erectus
|
| Binomial name | |
| †Homo erectus (Dubois, 1893)
| |
| Synonyms | |
Homo erectus (/ˌhoʊmoʊ əˈrɛktəs/ lit. 'upright man') is an extinct species of archaic human from the Pleistocene, with its earliest occurrence about 2 million years ago.[2] Its specimens are among the first recognizable members of the genus Homo.
Several human species, such as H. heidelbergensis and H. antecessor, appear to have evolved from H. erectus, and Neanderthals, Denisovans, and modern humans are in turn generally considered to have evolved from H. heidelbergensis.[3] H. erectus was the first human ancestor to spread throughout Eurasia, with a continental range extending from the Iberian Peninsula to Java. Asian populations of H. erectus may be ancestral to H. floresiensis[4] and possibly to H. luzonensis.[5] The last known population of H. erectus is H. e. soloensis from Java, around 117,000–108,000 years ago.[1]
H. erectus had a more modern gait and body proportions, and was the first human species to have exhibited a flat face, prominent nose, and possibly sparse body hair coverage. Though the species' brain size certainly exceeds that of ancestor species, capacity varied widely depending on the population. In earlier populations, brain development seemed to cease early in childhood, suggesting that offspring matured faster, thus limiting cognitive development. H. erectus was an apex predator;[6] sites generally show consumption of medium to large animals, such as bovines or elephants, and suggest the development of predatory behavior and coordinated hunting. H. erectus is associated with the Acheulean stone tool industry, and is postulated to have been the earliest human ancestor capable of using fire,[7] hunting and gathering in coordinated groups, caring for injured or sick group members, and possibly seafaring and art (though examples of art are controversial, and are otherwise rudimentary and few and far between). Intentional seafaring has also been a controversial claim.[8]
H. erectus males and females may have been roughly the same size as each other (i.e. exhibited reduced sexual dimorphism), which could indicate monogamy in line with general trends exhibited in primates. Size, nonetheless, ranged widely from 146–185 cm (4 ft 9 in – 6 ft 1 in) in height and 40–68 kg (88–150 lb) in weight. It is unclear if H. erectus was anatomically capable of speech, though it is postulated they communicated using some proto-language.
Taxonomy
[edit]Naming
[edit]
Contrary to the view Charles Darwin expressed in his 1871 book Descent of Man, many late-19th century evolutionary naturalists postulated that Asia, not Africa, was the birthplace of humankind as it is midway between Europe and America, providing optimal dispersal routes throughout the world (the Out of Asia theory). Among these was German naturalist Ernst Haeckel, who argued that the first human species evolved on the now-disproven hypothetical continent "Lemuria" in what is now Southeast Asia, from a species he termed "Pithecanthropus alalus" ("speechless apeman").[9] "Lemuria" had supposedly sunk below the Indian Ocean, so no fossils could be found to prove this. Nevertheless, Haeckel's model inspired Dutch scientist Eugène Dubois to journey to the Dutch East Indies. Because no directed expedition had ever discovered human fossils (the few known had all been discovered by accident), and the economy was strained by the Long Depression, the Dutch government refused to fund Dubois. In 1887, he enlisted in the Dutch East India Army as a medical officer, and was able to secure a post in 1887 in the Indies to search for his "missing link" in his spare time.[10] On Java, he found a skullcap in 1891 and a femur in 1892 (Java Man) dating to the late Pliocene or early Pleistocene at the Trinil site along the Solo River, which he named Pithecanthropus erectus ("upright apeman") in 1893. He attempted unsuccessfully to convince the European scientific community that he had found an upright-walking ape-man. Given few fossils of ancient humans had even been discovered at the time, they largely dismissed his findings as a malformed non-human ape.[11]
The significance of these fossils would not be realized until the 1927 discovery of what Canadian paleoanthropologist Davidson Black called "Sinanthropus pekinensis" (Peking Man) at the Zhoukoudian cave near Beijing, China. Black lobbied across North America and Europe for funding to continue excavating the site,[12] which has since become the most productive H. erectus site in the world.[13] Continued interest in Java led to further H. erectus fossil discoveries at Ngandong (Solo Man) in 1931, Mojokerto (Java Man) in 1936, and Sangiran (Java Man) in 1937. The Sangiran site yielded the best preserved Java Man skull.[14] German paleoanthropologist Franz Weidenreich provided much of the detailed description of the Chinese specimens in several monographs. The original specimens were lost during the Second Sino-Japanese War after an attempt to smuggle them out of China for safekeeping. Only casts remain.
Similarities between Java Man and Peking Man led Ernst Mayr to rename both as Homo erectus in 1950. Throughout much of the 20th century, anthropologists debated the role of H. erectus in human evolution. Early in the century, due in part to the discoveries at Java and Zhoukoudian, the belief that modern humans first evolved in Asia was widely accepted. A few naturalists—Charles Darwin the most prominent among them—theorized that humans' earliest ancestors were African. Darwin had pointed out that chimpanzees and gorillas, humans' closest relatives, evolved and exist only in Africa.[15] Darwin did not include orangutans among the great apes of the Old World, likely because he thought of orangutans as primitive humans rather than apes.[16] While Darwin considered Africa as the most probable birthplace of human ancestors, he also made the following statement about the geographic location of human origins in his book The Descent of Man, and Selection in Relation to Sex: "... it is useless to speculate on this subject; for two or three anthropomorphous apes, one the Dryopithecus …, existed in Europe during the Miocene age; and since so remote a period the earth has certainly undergone many great revolutions, and there has been ample time for migration on the largest scale." (1889, pp. 155–156).
In 1949, the species was reported in Swartkrans Cave, South Africa, by South African paleoanthropologists Robert Broom and John Talbot Robinson, who described it as "Telanthropus capensis".[17] Homo fossils have also been reported from nearby caves, but their species designation has been a tumultuous discussion. A few North African sites have additionally yielded H. erectus remains, which at first were classified as "Atlantanthropus mauritanicus" in 1951.[18] Beginning in the 1970s, propelled most notably by Richard Leakey, more were being unearthed in East Africa predominantly at the Koobi Fora site, Kenya, and Olduvai Gorge, Tanzania.[19]
Archaic human fossils unearthed across Europe used to be assigned to H. erectus, but have since been separated as H. heidelbergensis as a result of British physical anthropologist Chris Stringer's work.[20]
Evolution
[edit]−10 — – −9 — – −8 — – −7 — – −6 — – −5 — – −4 — – −3 — – −2 — – −1 — – 0 — |
| |||||||||||||||||||||||||||||

It has been proposed that H. erectus evolved from H. habilis about 2 Mya, though this has been called into question because they coexisted for at least a half a million years. Alternatively, a group of H. habilis may have been reproductively isolated, and only this group developed into H. erectus (cladogenesis).[21]
Because the earliest remains of H. erectus are found in both Africa and East Asia (in China as early as 2.1 Mya,[22][23][24] in South Africa 2.04 Mya[2][25]), it is debated where H. erectus evolved. A 2011 study suggested that it was H. habilis who reached West Asia from Africa, that early H. erectus developed there, and that early H. erectus would then have dispersed from West Asia to East Asia (Peking Man), Southeast Asia (Java Man), back to Africa (Homo ergaster), and to Europe (Tautavel Man), eventually evolving into modern humans in Africa.[26][27] Others have suggested that H. erectus/H. ergaster developed in Africa, where it eventually evolved into modern humans.[28][29]
H. erectus had reached Sangiran, Java, by 1.5 Mya,[30] and a second and distinct wave of H. erectus had colonized Zhoukoudian, China, about 780 kya (thousand years ago). Early teeth from Sangiran are bigger and more similar to those of basal (ancestral) Western H. erectus and H. habilis than to those of the derived Zhoukoudian H. erectus. However, later Sangiran teeth seem to reduce in size, which could indicate a secondary colonization event of Java by the Zhoukoudian or some closely related population.[31]
Subspecies
[edit]"Wushan Man" was proposed as Homo erectus wushanensis, but is now thought to be based upon fossilized fragments of an extinct non-hominin ape.[32]
Since the discovery of Java Man in 1893, there has been a trend in paleoanthropology of reducing the number of proposed species of Homo, to the point where H. erectus includes all early (Lower Paleolithic) forms of Homo sufficiently derived from H. habilis and distinct from early H. heidelbergensis (in Africa also known as H. rhodesiensis).[33] It is sometimes considered as a wide-ranging, polymorphous species.[34]
Due to such a wide range of variation, it has been suggested that the ancient H. rudolfensis and H. habilis should be considered early varieties of H. erectus.[35][36] The primitive H. e. georgicus from Dmanisi, Georgia has the smallest brain capacity of any known Pleistocene hominin (about 600 cc), and its inclusion in the species would greatly expand the range of variation of H. erectus to perhaps include species as H. rudolfensis, H. gautengensis, H. ergaster, and perhaps H. habilis.[37] However, a 2015 study suggested that H. georgicus represents an earlier, more primitive species of Homo derived from an older dispersal of hominins from Africa, with H. ergaster/erectus possibly deriving from a later dispersal.[38] H. georgicus is sometimes not even regarded as H. erectus.[39][40]
It is debated whether the African H. e. ergaster is a separate species (and that H. erectus evolved in Asia, then migrated to Africa),[41] or is the African form (sensu lato) of H. erectus (sensu stricto). In the latter, H. ergaster has also been suggested to represent the immediate ancestor of H. erectus.[42] It has also been suggested that H. ergaster instead of H. erectus, or some hybrid between the two, was the immediate ancestor of other archaic humans and modern humans.[citation needed] It has been proposed that Asian H. erectus have several unique characteristics from non-Asian populations (autapomorphies), but there is no clear consensus on what these characteristics are or if they are indeed limited to only Asia. Based on supposed derived characteristics, the 120 kya Javan H. e. soloensis has been proposed to have speciated from H. erectus, as H. soloensis, but this has been challenged because most of the basic cranial features are maintained.[43] In a wider sense, H. erectus had mostly been replaced by H. heidelbergensis by about 300 kya, with possible late survival of H. erectus soloensis in Java an estimated 117-108 kya.[1]
- H. e. bilzingslebenensis (Vlček 1978): Originally described from a series of skulls from Bilzingsleben, with the individual of Vertesszöllös being referred.[44] The material historically referred to this taxon are now affiliated with Neanderthals and the hominins at Sima de los Huesos.[45]
- H. e. capensis (Broom 1917): A variant of "Homo capensis",[46] a taxon erected from a skull from South Africa formally classified as a type of "race" but is now considered a representative of the Khoisan.[47]
- H. e. chenchiawoensis: A name utilized in a 2007 review of Chinese archeology; the text suggests that it and gongwanglingensis are contenders in taxonomy[48] (despite this name not appearing in the literature).
- H. e. erectus (Dubois 1891):[49] The Javanese specimens of H. erectus were once classified as a distinct subspecies in the 1970s. The cranium from Trinil is the holotype.[50]
- H. e. ergaster (Groves and Mazák 1975): Antón and Middleton (2023) suggested that ergaster should be disused based on poor diagnoses.[51] The name Homo erectus ergaster georgicus was created to classify the Dmanisi population as a subspecies of H. e. ergaster, but quadrinomials are not supported by the ICZN.[52]
- H. e. georgicus (Gabounia 1991):[53] This hypothetical subspecific designation unites the D2600 cranium with the remainder of the Dmanisi sample, a connection that was, at the time, controversial and was only suggested if the single-species hypothesis could be proven true.[54]
- H. e. gongwanglingensis: A name utilized in a 2007 review of Chinese archeology; the text suggests that it and chenchiawoensis are contenders in taxonomy.[48] Rukang (1992) notes that this taxon was born in a "subspecies fever".[55]
- H. e. habilis (Leakey, Tobias, and Napier 1964): D.R. Hughes believed that the Olduvai specimens were not distinct enough to be assigned to Australopithecus, so he created this taxon, as an early variation of H. erectus.[56]
- H. e. heidelbergensis (Schoetensack 1908): This taxon was used as an alternative to standard H. heidelbergensis during the middle 20th century, interpreted as a European chronospecies of the wider Middle Pleistocene hominin morph.[57]
- H. e. hexianensis (Huang 1982): Established based on the Hexian cranium.[58]
- H. e. hungaricus (Naddeo 2023): A Hungarian paper submitted to a conference lists this subspecies as an alternate name for the Vertesszöllös remains.[59]
- H. e. lantianensis (Ju-Kang 1964): Based on hominin fossils discovered in Lantian, originally named as a species of Sinanthropus and then reclassified as a subspecies.[60]
- H. e. leakeyi (Heberer 1963): A conditional name and thus unavailable for taxonomic use, once used to describe OH 9. The replacement name is louisleakeyi.[61] It received limited use as a subspecies.[46]
- H. e. mapaensis (Kurth 1965): A name that was proposed for the Maba cranium, although the use of the word 'perhaps' was interpreted by the Paleo Core database to be a conditional proposal and thus not available for valid reuse under the ICZN. Groves (1989) classified it as a subspecies of Homo sapiens, and Howell (1999) did not assign the species to a genus.[62]
- H. e. mauritanicus (Arambourg 1954): A subspecies that received limited use as a descriptor for the cranial and mandibular material discovered at Tighenif.[46]
- H. e. narmadensis (Sonakia 1984): The name given to the Narmada cranium.[63]
- H. e. newyorkensis (Laitman and Tattersall 2001): A name based on the Sambungmacan 3 cranium.[64]
- H. e. ngandongensis (Sartono 1976): A name that was used in the process of splitting Pithecanthropus into many subspecies.[65]
- H. e. olduvaiensis: A subspecies that described the OH 9 cranium, compared to the Bilzingsleben cranial fragments.[66]
- H. e. pekinensis (Black and Zdansky 1927): Originally assigned the type of Sinanthropus based on a single molar.[67] Antón and Middleton (2023) suggested that Zhoukoudian and Nanjing may be referrable under this name if they exhibit enough discontinuity from H. erectus proper.[51]
- H. e. reilingensis (Czarnetzki 1989): Referring to a single cranial fragment, this subspecies is now considered a member of the Neanderthal lineage.[68]
- H. e. soloensis (Oppenoorth 1932): The original name devised by Oppenoorth for the Ngandong crania.[69]
- H. e. tautavelensis (de Lumley and de Lumley 1971): Referring to the remains discovered at Arago, with many preferring allocation to Homo heidelbergensis.[70] The remains were determined not to be H. erectus by Antón and Middleton (2023).[51]
- H. e. trinilensis (Sartono 1976): A tentative classification scheme, thus making the name conditional and unable for use.[71]
- H. e. wadjakensis (Dubois 1921): A species established by Eugene Dubois based on the Wajak skulls.[72] Pramujiono classified these materials as a subspecies, and incorrectly self-published the name as wajakensis.[73]
- H. e. wushanensis (Huang and Fang 1991): Originally conceived as a hominin, the remains this taxon is founded on are more likely referred to Ponginae.[74][75]
- H. e. yuanmouensis (Li et al. 1977): Based on hominin remains[76] that Antón and Middleton (2023) suggest do not belong to the taxon H. erectus, although they do not provide an alternate classification.[51]

Descendants and synonyms
[edit]This section needs additional citations for verification. (July 2021) |
Homo erectus is the most long-lived species of Homo, having survived for almost two million years. By contrast, Homo sapiens emerged about a third of a million years ago.
Regarding many archaic humans, there is no definite consensus as to whether they should be classified as subspecies of H. erectus or H. sapiens or as separate species.
- African H. erectus candidates
- Homo ergaster (or "African H. erectus")
- Homo naledi
- Eurasian H. erectus candidates:
- Homo floresiensis[77]
- Homo rhodesiensis
- the Narmada fossil, discovered in 1982 in Madhya Pradesh, India, was at first suggested as H. erectus or Homo erectus narmadensis.[78]
Meganthropus, based on fossils found in Java, dated to between 1.4 and 0.9 Mya, was tentatively grouped with H. erectus in contrast to earlier interpretations of it as a giant species of early human[33] although older literature has placed the fossils outside of Homo altogether.[79] However, Zanolli et al. (2019) judged Meganthropus to be a distinct genus of extinct ape.[80]
Anatomy
[edit]Head
[edit]
Homo erectus featured a flat face compared to earlier hominins; pronounced brow ridge; and a low, flat skull.[81][82] The presence of sagittal, frontal, and coronal keels, which are small crests that run along these suture lines, has been proposed to be evidence of significant thickening of the skull, specifically the cranial vault. CT scan analyses reveal this to not be the case. However, the squamous part of occipital bone, particularly the internal occipital crest, at the rear of the skull is notably thicker than that of modern humans, likely a basal (ancestral) trait.[82][83] The fossil record indicates that H. erectus was the first human species to have featured a projecting nose, which is generally thought to have evolved in response to breathing dry air in order to retain moisture.[84] American psychologist Lucia Jacobs hypothesized that the projecting nose instead allowed for distinguishing the direction different smells come from (stereo olfaction) to facilitate navigation and long-distance migration.[85]
The average brain size of Asian H. erectus is about 1,000 cc (61 cu in). However, markedly smaller specimens have been found in Dmanisi, Georgia (H. e. georgicus); Koobi Fora and Olorgesailie, Kenya; and possibly Gona, Ethiopia. Overall, H. erectus brain size varies from 546–1,251 cc (33.3–76.3 cu in),[86] which is greater than the range of variation seen in modern humans and chimps, though less than that of gorillas.[citation needed]

In an article published in 2021 titled "Interpopulational variation in human brain size: Implications for hominin cognitive phylogeny," it was found that the brain size of Asian H. erectus over the last 600,000 years overlaps significantly with modern human populations. Significantly, some small brained modern populations showed greater affinity with H. erectus than they did with other large brained and large bodied modern populations. The paper points out methodological flaws in current understanding of brain size increase in human evolution, where species averages are compared with fossils, which overlooks interpopulational variation. It also overlooks the fact that some modern populations have not seen any dramatic brain size increase relative to H. erectus with most of the increase occurring in northern populations, which has the result of obscuring interpopulational variation. As the authors write '...the increase in the mean of H. sapiens cranial capacity is to a large extent due to an increase in the upper limit with a much less pronounced increase in the lower limit relative to our H. erectus sample. And this increase in the upper limit seems to be more pronounced in northern populations – which may be a result of correlated increases in body size in addition to climatic factors. Consequently, the authors argue that purely based on brain size similarities, Asian H. erectus could be re-classified as a subspecies of H. sapiens, that is H. sapiens soloensis - as was suggested by earlier authors.[87]
Dentally, H. erectus have the thinnest enamel of any Plio–Pleistocene hominin. Enamel prevents the tooth from breaking from hard foods but impedes shearing through tough foods. The bodies of the mandibles of H. erectus, and all early Homo, are thicker than those of modern humans and all living apes. The mandibular body resists torsion from the bite force or chewing, meaning their jaws could produce unusually powerful stresses while eating, but the practical application of this is unclear. Nonetheless, the mandibular bodies of H. erectus are somewhat thinner than those of early Homo. The premolars and molars also have a higher frequency of pits than H. habilis, suggesting H. erectus ate more brittle foods (which cause pitting). These all indicate that the H. erectus mouth was less capable of processing hard foods and more at shearing through tougher foods, thus reducing the variety of foods it could process, likely as a response to tool use.[88]
Body
[edit]
Like modern humans, H. erectus varied widely in size, ranging from 146–185 cm (4 ft 9 in – 6 ft 1 in) in height and 40–68 kg (88–150 lb) in weight, thought to be due to regional differences in climate, mortality rates, or nutrition.[89][90] Among primates, this marked of a response to environmental stressors (phenotypic plasticity) is only demonstrated in modern humans.[91][92][93]
Like modern humans and unlike other great apes, there does not seem to have been a great size disparity between H. erectus males and females (size-specific sexual dimorphism), though there is not much fossil data regarding this.[94] Brain size in two adults from Koobi Fora measured 848 and 804 cc (51.7 and 49.1 cu in),[86] and another significantly smaller adult measured 691 cc (42.2 cu in), which could possibly indicate sexual dimorphism, though sex was undetermined.[21] Another case that depicts the difficulty of assigning sex to the fossil record is a few samples taken in Olduvai Gorge. In 1960, in Olduvai Gorge two skulls identified as OH12 and OH9, were found to be that of H. erectus with a cranial capacities of 1000 cc and 700 cc.[95] It is unclear if sexual dimorphism is at play here since the remains are fragmentary.[95] If H. erectus did not exhibit sexual dimorphism, then it is possible that they were the first in the human line to do so, though the fragmentary fossil record for earlier species makes this unclear. If yes, then there was a substantial and sudden increase in female height.[96] Certain features of sexual dimorphism are often identified in the possibility of determining sex such as lack of muscle marking.[97]

H. erectus had about the same limb configurations and proportions as modern humans, implying humanlike locomotion,[98] the first in the Homo lineage.[91] H. erectus tracks near Ileret, Kenya, also indicate a human gait.[99] A humanlike shoulder suggests an ability for high speed throwing.[100] It was once thought that Turkana boy had 6 lumbar vertebra instead of the 5 seen in modern humans and 11 instead of 12 thoracic vertebrae, but this has since been revised, and the specimen is now considered to have exhibited a humanlike curvature of the spine (lordosis) and the same number of respective vertebrae.[101]
It is largely unclear when human ancestors lost most of their body hair. Genetic analysis suggests that high activity in the melanocortin 1 receptor, which would produce dark skin, dates back to 1.2 Mya. This could indicate the evolution of hairlessness around this time, as a lack of body hair would have left the skin exposed to harmful UV radiation.[102] It is possible that exposed skin only became maladaptive in the Pleistocene, because the increasing tilt of the Earth (which also caused the ice ages) would have increased solar radiation bombardment- which would suggest that hairlessness first emerged in the australopithecines.[103] However, australopithecines seem to have lived at much higher, much colder elevations—typically 1,000–1,600 m (3,300–5,200 ft) where the nighttime temperature can drop to 10 or 5 °C (50 or 41 °F)—so they may have required hair to stay warm, unlike early Homo which inhabited lower, hotter elevations.[104] Populations in higher latitudes potentially developed lighter skin to prevent vitamin D deficiency.[105] A 500–300 kya H. erectus specimen from Turkey was diagnosed with the earliest known case of tuberculous meningitis, which is typically exacerbated in dark-skinned people living in higher latitudes due to vitamin D deficiency.[106] Hairlessness is generally thought to have facilitated sweating,[107] but reduction of parasite load and sexual selection have also been proposed.[108][109]
Metabolism
[edit]
The 1.8 Ma Mojokerto child specimen from Java, who died at about 1 year of age, presented 72–84% of the average adult brain size, which is more similar to the faster brain growth trajectory of great apes than modern humans. This indicates that H. erectus was probably not cognitively comparable to modern humans, and that secondary altriciality—an extended childhood and long period of dependency due to the great amount of time required for brain maturation—evolved much later in human evolution, perhaps in the modern human/Neanderthal last common ancestor.[110] It was previously believed that, based on the narrow pelvis of Turkana boy, H. erectus could only safely deliver a baby with a brain volume of about 230 cc (14 cu in), equating to a similar brain growth rate as modern humans to achieve the average adult brain size of 600–1,067 cc (36.6–65.1 cu in). However, a 1.8 Ma female pelvis from Gona, Ethiopia, shows that H. erectus babies with a brain volume of 310 cc (19 cu in) could have been safely delivered, which is 34–36% the mean adult size, compared to 40% in chimps and 28% in modern humans. This more aligns with the conclusions drawn from the Mojokerto child.[94] A faster development rate could indicate a lower expected lifespan.[111]
Based on an average mass of 63 kg (139 lb) for males and 52.3 kg (115 lb) for females, the daily energy expenditure (DEE)—the amount of calories metabolized in one day—was estimated to be about 2271.8 and 1909.5 kcal, respectively. This is similar to that of earlier Homo, despite a marked increase in activity and migratory capacity, likely because the longer legs of H. erectus were more energy-efficient in long-distance movement. Nonetheless, the estimate for H. erectus females is 84% higher than that for Australopithecus females, possibly due to an increased body size and a decreased growth rate.[112] A 2011 study, assuming high energy or dietary fat requirements based on the abundance of large game animals at H. erectus sites, calculated a DEE of 2,700–3,400 kcal of which 27–44% derived from fat, and 44–62% of the fat from animal sources. In comparison, modern humans with a similar activity level have a DEE of 2,450 calories, of which 33% derives from fat, and 49% of the fat from animals.[113]
Bone thickness
[edit]
The cortical bone (the outer layer of the bone) is extraordinarily thickened, particularly in East Asian populations. The skullcaps have oftentimes been confused with fossil turtle carapaces,[114] and the medullary canal in the long bones (where the bone marrow is stored, in the limbs) is extremely narrowed (medullary stenosis). This degree of thickening is usually exhibited in semi-aquatic animals which used their heavy (pachyosteosclerotic) bones as ballasts to help them sink, induced by hypothyroidism. Male specimens have thicker cortical bone than females.[115]
It is largely unclear what function this could have served. All pathological inducers would leave scarring or some other indicator not normally exhibited in H. erectus. Before more complete skeletons were discovered, Weidenreich suggested H. erectus was a gigantic species, thickened bone required to support the massive weight. It was hypothesized that intense physical activity could have induced bone thickening, but in 1970, human biologist Stanley Marion Garn demonstrated there is a low correlation between the two at least in modern humans. Garn instead noted different races have different average cortical bone thicknesses, and concluded it is genetic rather than environmental. It is unclear if the condition is caused by increased bone apposition (bone formation) or decreased bone resorption, but Garn noted the stenosis is quite similar to the congenital condition in modern humans induced by hyper-apposition. In 1985, biological anthropologist Gail Kennedy argued for resorption as a result of hyperparathyroidism caused by hypocalcemia (calcium deficiency), a consequence of a dietary shift to low-calcium meat. Kennedy could not explain why the calcium metabolism of H. erectus never adjusted.[115] In 1985, American paleoanthropologist Mary Doria Russell and colleagues argued the supraorbital torus is a response to withstanding major bending moment which localizes in that region when significant force is applied through the front teeth, such as while using the mouth as a third hand to carry objects.[116]
In 2004, Noel Boaz and Russel Ciochon suggested it was a result of a cultural practice, wherein H. erectus would fight each other with fists, stones, or clubs to settle disputes or battle for mates, since the skull is reinforced in key areas. The mandible is quite robust, capable of absorbing heavy blows (no "glass jaw"); the heavy brow ridge protects the eyes, and transitions into a bar covering the ears, connecting all the way in the back of the skull, meaning blows to any of these regions can be effectively dissipated across the skull; and the sagittal keel protects the top of the braincase. Many skullcaps bear usually debilitating fractures, such as the Peking Man skull X, yet they can show signs of surviving and healing. Anthropologist Peter Brown suggested a similar reason for the unusual thickening of the modern Australian Aboriginal skull, a result of a ritual popular in central and southeast Australian tribes where adversaries would wack each other with waddies (sticks) until knockout.[114]
Culture
[edit]Social structure
[edit]
The only fossil evidence regarding H. erectus group composition comes from four sites outside of Ileret, Kenya, where 97 footprints made 1.5 Mya were likely left by a group of at least 20 individuals. One of these trackways, based on the size of the footprints, may have been an entirely male group, which could indicate they were some specialised task group, such as a hunting or foraging party, or a border patrol. If correct, this would also indicate sexual division of labour, which distinguishes human societies from those of other great apes and social mammalian carnivores. In modern hunter gatherer societies who target large prey items, typically male parties are dispatched to bring down these high-risk animals, and, due to the low success rate, female parties focus on more predictable foods.[99] Based on modern day savanna chimp and baboon group composition and behavior, H. erectus ergaster may have lived in large, multi-male groups in order to defend against large savanna predators in the open and exposed environment.[117] However, dispersal patterns indicate that H. erectus generally avoided areas with high carnivore density.[118] It is possible that male–male bonding and male–female friendships were important societal aspects.[117]
Because H. erectus children had faster brain growth rates, H. erectus likely did not exhibit the same degree of maternal investment or child-rearing behaviours as modern humans.[94]
Because H. erectus males and females are thought to have been about the same size compared to other great apes (exhibit less size-specific sexual dimorphism), it is generally hypothesised that they lived in a monogamous society, as reduced sexual dimorphism in primates is typically correlated with this mating system.[96] However, it is unclear if H. erectus did in fact exhibit humanlike rates of sexual dimorphism.[21] If they did, then it would mean only female height increased from the ancestor species, which could have been caused by a shift in female fertility or diet, and/or reduced pressure on males for large size. This in turn could imply a shift in female behavior which made it difficult for males to maintain a harem, and vice versa.[119]
Food
[edit]Increasing brain size is often directly associated with a meatier diet and resultant higher caloric intake. Human entomophagy and therefore an increase in protein consumption through insects has also been proposed as a possible cause. However, it is also possible that the energy-expensive guts decreased in size in H. erectus, because the large ape gut is used to synthesize fat by fermenting plant matter, which was replaced by dietary animal fat, allowing more energy to be diverted to brain growth. This would have increased brain size indirectly while maintaining the same caloric requirements of ancestor species. H. erectus may have also been the first to use a hunting and gathering food collecting strategy as a response to the increasing dependence on meat. With an emphasis on teamwork, division of labor, and food sharing, hunting and gathering was a dramatically different subsistence strategy from previous modes.[88][113]

H. erectus sites frequently are associated with assemblages of medium- to large-sized game, namely elephants, rhinos, hippos, bovines, and boars. H. erectus would have had considerable leftovers, potentially pointing to food sharing or long-term food preservation (such as by drying) if most of the kill was indeed utilized. It is possible that H. erectus grew to become quite dependent on large-animal meat, and the disappearance of H. erectus from the Levant is correlated with the local extinction of the straight-tusked elephant.[113] Nonetheless, H. erectus diet likely varied widely depending upon location. For example, at the 780 kya Gesher Benot Ya'aqov site, Israel, the inhabitants gathered and ate 55 different types of fruits, vegetables, seeds, nuts, and tubers, and it appears that they used fire to roast certain plant materials that otherwise would have been inedible; they also consumed amphibians, reptiles, birds, aquatic and terrestrial invertebrates, in addition to the usual large creatures such as elephant and fallow deer.[120] At the 1.95 Mya FwJJ20 lakeside site in the East Turkana Basin, Kenya, the inhabitants ate (alongside the usual bovids, hippos, and rhinos) aquatic creatures such as turtles, crocodiles, and catfish. The large animals were likely scavenged at this site, but the turtles and fish were possibly collected live.[121] In East Africa between 2.0 and 1.4 Mya, carcasses of C4-grazing ungulates, particularly alcelaphins, featured increasingly prominently in the diet of these hominins.[122] At the 1.5 Mya Trinil H. K. site, Java, H. erectus likely gathered fish and shellfish.[123]
Dentally, H. erectus mouths were not as versatile as those of ancestor species, capable of processing a narrower range of foods. However, tools were likely used to process hard foods, thus affecting the chewing apparatus. This combination may have instead increased dietary flexibility (though this does not equate to a highly varied diet). Such versatility may have permitted H. erectus to inhabit a range of different environments and migrate beyond Africa.[88]
In 1999, British anthropologist Richard Wrangham proposed the "cooking hypothesis" which states that H. erectus speciated from the ancestral H. habilis because of fire usage and cooking 2 million years ago to explain the rapid doubling of brain size between these two species in only a 500,000-year timespan, and the sudden appearance of the typical human body plan. Cooking makes protein more easily digestible, speeds up nutrient absorption, and destroys food-borne pathogens, which would have increased the environment's natural carrying capacity, allowing group size to expand, causing selective pressure for sociality, requiring greater brain function.[124][125] However, the fossil record does not associate the emergence of H. erectus with fire usage nor with any technological breakthrough for that matter, and cooking likely did not become a common practice until after 400 kya.[88][113]
Java Man's dispersal through Southeast Asia coincides with the extirpation of the giant turtle Megalochelys, possibly due to overhunting as the turtle would have been an easy, slow-moving target which could have been stored for quite some time.[126]
Technology
[edit]Tool production
[edit]

H. erectus is credited with inventing the Acheulean stone tool industry, succeeding the Oldowan industry,[127][128] and were the first to make lithic flakes bigger than 10 cm (3.9 in), and hand axes (which includes bifacial tools with only 2 sides, such as picks, knives, and cleavers).[129] Though larger and heavier, these hand axes had sharper, chiseled edges.[130] They were likely multi-purpose tools, used in variety of activities such as cutting meat, wood, or edible plants.[131] In 1979, American paleontologist Thomas Wynn stated that Acheulean technology required operational intelligence (foresight and planning), being markedly more complex than Oldowan technology which included lithics of unstandardized shape, cross-sections, and symmetry. Based on this, he concluded that there is not a significant disparity in intelligence between H. erectus and modern humans and that, for the last 300,000 years, increasing intelligence has not been a major influencer of cultural evolution.[132] However, a 1 year old H. erectus specimen shows that this species lacked an extended childhood required for greater brain development, indicating lower cognitive capabilities.[110] A few sites, likely due to occupation over several generations, features hand axes en masse, such as at Melka Kunture, Ethiopia; Olorgesailie, Kenya; Isimila, Tanzania; and Kalambo Falls, Zambia.[131]
The earliest record of Acheulean technology comes from West Turkana, Kenya 1.76 Mya. Oldowan lithics are also known from the site, and the two seemed to coexist for some time. The earliest records of Acheulean technology outside of Africa date to no older than 1 Mya, indicating it only became widespread after some secondary H. erectus dispersal from Africa.[130]
On Java, H. erectus produced tools from shells at Sangiran[133] and Trinil.[134] Spherical stones, measuring 6–12 cm (2.4–4.7 in) in diameter, are frequently found in African and Chinese Lower Paleolithic sites, and were potentially used as bolas; if correct, this would indicate string and cordage technology.[135]
Fire
[edit]H. erectus is credited as the first human ancestor to have used fire, though the timing of this invention is debated mainly because campfires very rarely and very poorly preserve over long periods of time, especially thousands or millions of years. The earliest claimed fire sites are in Kenya, FxJj20 at Koobi Fora[136][124][137] and GnJi 1/6E in the Chemoigut Formation, as far back as 1.5 Mya,[124][137] and in South Africa, Wonderwerk Cave, 1.7 Mya.[138] The first firekeepers are thought to have simply transported to caves and maintained naturally occurring fires for extended periods of time or only sporadically when the opportunity arose. Maintaining fires would require firekeepers to have knowledge of slow-burning materials such as dung.[124] Fire becomes markedly more abundant in the wider archaeological record after 400,000–300,000 years ago, which can be explained as some advancement in fire management techniques took place at this time[124] or human ancestors only opportunistically used fire until this time.[137][139][88][113] It is possible that firestarting was invented and lost and reinvented multiple times and independently by different communities rather than being invented in one place and spreading throughout the world.[139] The earliest evidence of hearths comes from Gesher Benot Ya'aqov, Israel, over 700,000 years ago, where fire is recorded in multiple layers in an area close to water, both uncharacteristic of natural fires.[125]
Artificial lighting may have led to increased waking hours—modern humans have about a 16-hour waking period, whereas other apes are generally awake from only sunup to sundown—and these additional hours were probably used for socializing. Because of this, fire usage is probably also linked to the origin of language.[124][125] Artificial lighting may have also made sleeping on the ground instead of the trees possible by keeping terrestrial predators at bay.[125]
Migration into the frigid climate of Ice Age Europe may have only been possible because of fire, but evidence of fire usage in Europe until about 400–300,000 years ago is notably absent.[137] If these early European H. erectus did not have fire, it is largely unclear how they stayed warm, avoided predators, and prepared animal fat and meat for consumption. There was also a lower likelihood of naturally occurring fires due to lightning being less common in areas further north. It is possible that they only knew how to maintain fires in certain settings in the landscapes and prepared food some distance away from home, meaning evidence of fire and evidence of hominin activity are spaced far apart.[125] Alternatively, H. erectus may have only pushed farther north during warmer interglacial periods—thus not requiring fire, food storage, or clothing technology—[140] and their dispersal patterns indicate they generally stayed in warmer lower-to-middle latitudes.[118] It is debated if the H. e. pekinensis inhabitants of Zhoukoudian, Northern China, were capable of controlling fires as early as 770 kya to stay warm in what may have been a relatively cold climate.[141]
Construction
[edit]
In 1962, a 366 cm × 427 cm × 30 cm (12 ft × 14 ft × 1 ft) circle made with volcanic rocks was discovered in Olduvai Gorge. At 61–76 cm (2–2.5 ft) intervals, rocks were piled up to 15–23 cm (6–9 in) high. British palaeoanthropologist Mary Leakey suggested the rock piles were used to support poles stuck into the ground, possibly to support a windbreak or a rough hut. Some modern-day nomadic tribes build similar low-lying rock walls to build temporary shelters upon, bending upright branches as poles and using grasses or animal hide as a screen.[143] Dating to 1.75 Mya, it is the oldest claimed evidence of architecture.[144]
The earliest evidence of cave habitation is Wonderwerk Cave, South Africa, about 1.6 Mya, but evidence of cave use globally is sporadic until about 600 kya.[145]
Clothing
[edit]_Fundort_Nariokotome,_Kenia,_Rekonstruktion_im_Neanderthal_Museum.jpg)
It is largely unclear when clothing was invented, with the earliest estimate stretching as far back as 3 Mya to compensate for a lack of insulating body hair.[103] It is known that head lice and body lice (the latter can only inhabit clothed individuals) for modern humans diverged about 170 kya, well before modern humans left Africa, meaning clothes were already well in use before encountering cold climates. One of the first uses of animal hide is thought to have been for clothing, and the oldest hide scrapers date to about 780 kya; however, this is not indicative of clothing.[146]
Possible seafaring
[edit]Acheulean artifacts discovered on isolated islands that were never connected to land in the Pleistocene may show seafaring by H. erectus as early as 1 Mya in Indonesia. They had arrived on the islands of Flores, Timor, and Roti, which would have necessitated crossing the Lombok Strait (the Wallace Line), at least before 800 kya. It is also possible they were the first European mariners as well and crossed the Strait of Gibraltar between North Africa and Spain. A 2021 genetic analysis of these island populations of H. erectus found no evidence of interbreeding with modern humans.[147] Seafaring capability would show H. erectus had a great capacity for planning, likely months in advance of the trip.[148][149] However, there is no evidence of the watercrafts required for such seafaring, though these would likely not survive in the fossil record. Additionally, experiments by Bednarik that show modern humans can make complex boats using materials available to H. erectus, actually undercut this hypothesis. Such experiments presuppose cognition rivalling H. sapiens, despite the lack of evidence for such a premise. Instead, it may well be that Homo erectus reached Flores through natural rafts of vegetation that had been swept out to sea following a cyclone or tsunami.[8]
Similarly, Homo luzonensis is dated between 771,000 and 631,000 years ago. Because Luzon has always been an island in the Quaternary, the ancestors of H. luzonensis would have had to have made a substantial sea crossing and crossed the Huxley Line.[150]
Healthcare
[edit].jpg)
The earliest probable example of infirming sick group members is a 1.77 Mya H. e. georgicus specimen who had lost all but one tooth due to age or gum disease, the earliest example of severe chewing impairment, yet still survived for several years afterwards. However, it is possible australopithecines were capable of caring for debilitated group members.[151] Unable to chew, this H. e. georgicus individual probably ate soft plant or animal foods possibly with assistance from other group members. High-latitude groups are thought to have been predominantly carnivorous, eating soft tissue such as bone marrow or brains, which may have increased survival rates for toothless individuals.[152]
The 1.5 Mya Turkana boy was diagnosed with juvenile spinal disc herniation, and, because this specimen was still growing, this caused some scoliosis (abnormal curving of the spine). These usually cause recurrent lower back pain and sciatica (pain running down the leg), and likely restricted Turkana boy in walking, bending, and other daily activities. The specimen appears to have survived into adolescence, which evidences advanced group care.[153]
The 1,000–700 kya Java man specimen presents a noticeable osteocyte on the femur, likely Paget's disease of bone, and osteopetrosis, thickening of the bone, likely resulting from skeletal fluorosis caused by ingestion of food contaminated by fluorine-filled volcanic ash (as the specimen was found in ash-filled strata). Livestock that grazes on volcanic ash ridden fields typically die of acute intoxication within a few days or weeks.[154]
Art and rituals
[edit].jpg)

An engraved Pseudodon shell DUB1006-fL with geometric markings could possibly be evidence of the earliest art-making, dating back to 546–436 kya. Art-making capabilities could be considered evidence of symbolic thinking, which is associated with modern cognition and behavior.[134][155][156][157] In 1976, American archeologist Alexander Marshack asserted that engraved lines on an ox rib, associated with Acheulean lithics, from Pech de l'Azé, France, are similar to a meander design found in modern human Upper Paleolithic cave art.[158] Three ostrich eggshell beads associated with Achuelian lithics were found in northwestern Africa, the earliest disc beads ever found, and Acheulian disc beads have also been found in France and Israel.[148] The Middle Pleistocene "Venus of Tan-Tan" and "Venus of Berekhat Ram" are postulated to been crafted by H. erectus to resemble a human form. They were mostly formed by natural weathering, but slightly modified to emphasize certain grooves to suggest hairline, limbs, and eyes.[159][160] The former has traces of pigments on the front side, possibly indicating it was colored.[159]
H. erectus was also the earliest human to have intentionally collected red-colored pigments, namely ochre, recorded as early as the Middle Pleistocene. Ochre lumps at Olduvai Gorge, Tanzania—associated with the 1.4 Ma Olduvai Hominid 9—and Ambrona, Spain—which dates to 424–374 kya—were suggested to have been struck by a hammerstone and purposefully shaped and trimmed.[161][158] At Terra Amata, France—which dates to 425–400 or 355–325 kya—red, yellow, and brown ochres were recovered in association with pole structures; ochre was probably heated to achieve such a wide color range.[161][162] As it is unclear if H. erectus could have used ochre for any practical application, ochre collection might indicate that H. erectus was the earliest human to have exhibited a sense of aesthetics and to think beyond simply survival. Later human species are postulated to have used ochre as body paint, but in the case of H. erectus, it is contested if body paint was used so early in time. Further, it is unclear if these few examples are not simply isolated incidents of ochre use, as ochre is much more prevalent in Middle and Upper Paleolithic sites attributed to Neanderthals and H. sapiens.[163][158]
In 1935, Jewish-German anthropologist Franz Weidenreich speculated that the inhabitants of the Chinese Zhoukoudian Peking Man site were members of some Lower Paleolithic Skull Cult because the skulls all showed fatal blows to the head, breaking in of the foramen magnum at the base of the skull, by-and-large lack of preserved facial aspects, an apparently consistent pattern of breaking on the mandible, and a lack of post-cranial remains (elements that are not the skull). He believed that the inhabitants were headhunters, and smashed open the skulls and ate the brains of their victims.[164][158] However, scavenging animals and natural forces such as flooding can also inflict the same kind of damage to skulls,[158] and there is not enough evidence to suggest manhunting or cannibalism.[165]
In 1999, British science writers Marek Kohn and Steven Mithen said that many hand axes exhibit no wear and were produced en masse, and concluded that these symmetrical, tear-drop shaped lithics functioned primarily as display tools so males could prove their fitness to females in some courting ritual, and were discarded afterwards.[166] However, an apparent lack of reported wearing is likely due to a lack of use-wear studies, and only a few sites yield an exorbitant sum of hand axes likely due to gradual accumulation over generations instead of mass production.[131]
Language
[edit]In 1984, the vertebral column of the 1.6 Mya adolescent Turkana boy indicated that this individual did not have properly developed respiratory muscles in order to produce speech. In 2001, American anthropologists Bruce Latimer and James Ohman concluded that Turkana boy was afflicted by skeletal dysplasia and scoliosis.[167] In 2006, American anthropologist Marc Meyer and colleagues described a 1.8 Mya H. e. georgicus specimen as having a spine within the range of variation of modern human spines, contending that Turkana boy had spinal stenosis and was thus not representative of the species. Also, because he considered H. e. georgicus ancestral to all non-African H. erectus, Meyer concluded that the respiratory muscles of all H. erectus (at least non-H. ergaster) would not have impeded vocalisation or speech production.[168] However, in 2013 and 2014, anthropologist Regula Schiess and colleagues concluded that there is no evidence of any congenital defects in Turkana boy, and considered the specimen representative of the species.[169][170]
Neurologically, all Homo have similarly configured brains, and, likewise, the Broca's and Wernicke's areas (in charge of sentence formulation and speech production in modern humans) of H. erectus were comparable to those of modern humans. However, this is not indicative of anything in terms of speech capability as even large chimpanzees can have similarly expanded Broca's area, and it is unclear if these areas served as language centers in archaic humans.[171] A 1-year-old H. erectus specimen shows that an extended childhood to allow for brain growth, which is a prerequisite in language acquisition, was not exhibited in this species.[110]
The hyoid bone supports the tongue and makes possible modulation of the vocal tract to control pitch and volume. A 400 kya H. erectus hyoid bone from Castel di Guido, Italy, is bar-shaped—more similar to that of other Homo than to that of non-human apes and Australopithecus—but is devoid of muscle impressions, has a shield-shaped body, and is implied to have had reduced greater horns, meaning H. erectus lacked a humanlike vocal apparatus and thus anatomical prerequisites for a modern human level of speech.[172] Increasing brain size and cultural complexity in tandem with technological refinement, and the hypothesis that articulate Neanderthals and modern humans may have inherited speech capabilities from the last common ancestor, could possibly indicate that H. erectus used some proto-language and built the basic framework which fully fledged languages would eventually be built around.[173] However, this ancestor may have instead been H. heidelbergensis, as a hyoid bone of a 530 kya H. heidelbergensis specimen from the Spanish Sima de los Huesos Cave is like that of modern humans,[174] and another specimen from the same area shows an auditory capacity sensitive enough to pick up human speech.[175]
Extinction
[edit]This section needs expansion. You can help by adding to it. (December 2020) |
The last known occurrence of Homo erectus is 117,000–108,000 years ago in Ngandong, Java according to a study published in 2019.[1]
In 2020, researchers reported that Homo erectus and Homo heidelbergensis lost more than half of their climate niche – climate they were adapted to – with no corresponding reduction in physical range, just before extinction and that climate change played a substantial role in extinctions of past Homo species.[176][177][178]
Fossils
[edit]
The lower cave of China's Zhoukoudian Cave is one of the most important archaeological sites worldwide.[179] There have been remains of 45 Homo erectus individuals found and thousands of tools recovered.[179] Most of these remains were lost during World War 2, with the exception of two postcranial elements that were rediscovered in China in 1951 and four human teeth from 'Dragon Bone Hill'.[179]
New evidence has shown that Homo erectus does not have uniquely thick vault bones, as was previously thought.[180] Testing showed that neither Asian nor African Homo erectus had uniquely large vault bones.[180]
Individual fossils
[edit]Some of the major Homo erectus fossils:
- Indonesia (island of Java): Trinil 2 (holotype), Sangiran collection, Sambungmachan collection,[181] Ngandong collection
- China ("Peking Man"): Lantian (Gongwangling and Chenjiawo), Yunxian, Zhoukoudian, Nanjing, Hexian
- Kenya: KNM ER 3883, KNM ER 3733
- Vietnam: Northern, Tham Khuyen,[182] Hoa Binh[citation needed]
- Republic of Georgia: Dmanisi collection ("Homo erectus georgicus")
- Ethiopia: Daka calvaria
- Eritrea: Buia cranium (possibly H. ergaster)[183]
- Denizli Province, Turkey: Kocabas fossil[106]
- Drimolen, South Africa: DNH 134[184]
Gallery
[edit]-
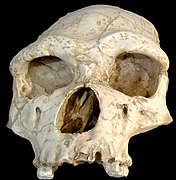 Homo erectus tautavelensis skull.
Homo erectus tautavelensis skull. -

-

-
 A reconstruction based on evidence from the Daka Member, Ethiopia
A reconstruction based on evidence from the Daka Member, Ethiopia -
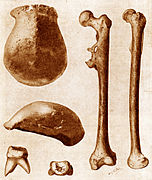 Original fossils of Pithecanthropus erectus (now Homo erectus) found in Java in 1891.
Original fossils of Pithecanthropus erectus (now Homo erectus) found in Java in 1891.


.jpg)


See also
[edit]General:
- List of fossil sites (with link directory)
- List of human evolution fossils (with images)
References
[edit]- ^ a b c d Rizal Y, Westaway KE, Zaim Y, van den Bergh GD, Bettis EA, Morwood MJ, et al. (January 2020). "Last appearance of Homo erectus at Ngandong, Java, 117,000-108,000 years ago". Nature. 577 (7790): 381–385. doi:10.1038/s41586-019-1863-2. PMID 31853068. S2CID 209410644.
- ^ a b Herries AI, Martin JM, Leece AB, Adams JW, Boschian G, Joannes-Boyau R, et al. (April 2020). "Contemporaneity of Australopithecus, Paranthropus, and early Homo erectus in South Africa". Science. 368 (6486): eaaw7293. doi:10.1126/science.aaw7293. PMID 32241925.
- ^ Dembo M, Radovčić D, Garvin HM, Laird MF, Schroeder L, Scott JE, et al. (August 2016). "The evolutionary relationships and age of Homo naledi: An assessment using dated Bayesian phylogenetic methods". Journal of Human Evolution. 97: 17–26. Bibcode:2016JHumE..97...17D. doi:10.1016/j.jhevol.2016.04.008. hdl:2164/8796. PMID 27457542.
- ^ van den Bergh GD, Kaifu Y, Kurniawan I, Kono RT, Brumm A, Setiyabudi E, et al. (June 2016). "Homo floresiensis-like fossils from the early Middle Pleistocene of Flores". Nature. 534 (7606): 245–248. Bibcode:2016Natur.534..245V. doi:10.1038/nature17999. PMID 27279221. S2CID 205249218.
- ^ Détroit F, Mijares AS, Corny J, Daver G, Zanolli C, Dizon E, et al. (April 2019). "A new species of Homo from the Late Pleistocene of the Philippines" (PDF). Nature. 568 (7751): 181–186. Bibcode:2019Natur.568..181D. doi:10.1038/s41586-019-1067-9. PMID 30971845. S2CID 106411053.
- ^ Ben-Dor M, Sirtoli R, Barkai R (August 2021). "The evolution of the human trophic level during the Pleistocene". American Journal of Physical Anthropology. 175 (Suppl 72): 27–56. doi:10.1002/ajpa.24247. PMID 33675083.
- ^ Zohar, Irit; Alperson-Afil, Nira; Goren-Inbar, Naama; Prévost, Marion; Tütken, Thomas; Sisma-Ventura, Guy; Hershkovitz, Israel; Najorka, Jens (14 November 2022). "Evidence for the cooking of fish 780,000 years ago at Gesher Benot Ya'aqov, Israel". Nature Ecology & Evolution. 6 (12): 2016–2028. Bibcode:2022NatEE...6.2016Z. doi:10.1038/s41559-022-01910-z. ISSN 2397-334X. PMID 36376603. S2CID 253522354.
- ^ a b Botha, Rudolf (16 April 2024). "Did Homo erectus Have Language? The Seafaring Inference". Cambridge Archaeological Journal: 1–17. doi:10.1017/S0959774324000118. ISSN 0959-7743.
- ^ Theunissen 2012, p. 6.
- ^ Theunissen 2012, p. 33.
- ^ Yen HP (2014). "Evolutionary Asiacentrism, Peking man, and the origins of sinocentric ethno-nationalism". Journal of the History of Biology. 47 (4): 585–625. doi:10.1007/s10739-014-9381-4. PMID 24771020. S2CID 23308894.
- ^ Sigmon 1981, p. 64.
- ^ Yang L (2014). "Zhoukoudian: Geography and Culture". Encyclopedia of Global Archaeology. Springer Science+Business Media. pp. 7961–7965. doi:10.1007/978-1-4419-0465-2_1899. ISBN 978-1-4419-0466-9.
- ^ Ciochon RL, Huffman OF (2014). "Java Man". In Smith C (ed.). Encyclopedia of Global Archaeology. pp. 4182–4188. doi:10.1007/978-1-4419-0465-2_712. ISBN 978-1-4419-0426-3. S2CID 241324984.
- ^ Darwin CR (1871). The Descent of Man and Selection in Relation to Sex. John Murray. ISBN 978-0-8014-2085-6.
- ^ van Wyhe, John; Kjærgaard, Peter C. (1 June 2015). "Going the whole orang: Darwin, Wallace and the natural history of orangutans". Studies in History and Philosophy of Science Part C: Studies in History and Philosophy of Biological and Biomedical Sciences. 51: 53–63. doi:10.1016/j.shpsc.2015.02.006. ISSN 1369-8486. PMID 25861859. S2CID 20089470.
- ^ Curnoe D (June 2010). "A review of early Homo in southern Africa focusing on cranial, mandibular and dental remains, with the description of a new species (Homo gautengensis sp. nov.)". Homo. 61 (3): 151–177. doi:10.1016/j.jchb.2010.04.002. PMID 20466364.
- ^ Sigmon 1981, p. 231.
- ^ Sigmon 1981, p. 193.
- ^ de Lumley MA (2015). "L'homme de Tautavel. Un Homo erectus européen évolué. Homo erectus tautavelensis" [Tautavel Man. An evolved European Homo erectus. Homo erectus tautavelensis]. L'Anthropologie (in French). 119 (3): 342–344. doi:10.1016/j.anthro.2015.06.001.
- ^ a b c Spoor F, Leakey MG, Gathogo PN, Brown FH, Antón SC, McDougall I, et al. (August 2007). "Implications of new early Homo fossils from Ileret, east of Lake Turkana, Kenya". Nature. 448 (7154): 688–691. Bibcode:2007Natur.448..688S. doi:10.1038/nature05986. PMID 17687323. S2CID 35845.
- ^ Zhu Z, Dennell R, Huang W, Wu Y, Qiu S, Yang S, et al. (July 2018). "Hominin occupation of the Chinese Loess Plateau since about 2.1 million years ago". Nature. 559 (7715): 608–612. Bibcode:2018Natur.559..608Z. doi:10.1038/s41586-018-0299-4. PMID 29995848. S2CID 49670311.
- ^ Barras C (2018). "Tools from China are oldest hint of human lineage outside Africa". Nature. doi:10.1038/d41586-018-05696-8. ISSN 0028-0836. S2CID 188286436.
- ^ Hao L, Chao Rong L, Kuman K (2017). "Longgudong, an Early Pleistocene site in Jianshi, South China, with stratigraphic association of human teeth and lithics". Science China Earth. 60 (3): 452–462. Bibcode:2017ScChD..60..452L. doi:10.1007/s11430-016-0181-1. S2CID 132479732.
- ^ "Our direct human ancestor Homo erectus is older than we thought". EurekAlert. AAAS.
- ^ Ferring R, Oms O, Agustí J, Berna F, Nioradze M, Shelia T, et al. (June 2011). "Earliest human occupations at Dmanisi (Georgian Caucasus) dated to 1.85-1.78 Ma". Proceedings of the National Academy of Sciences of the United States of America. 108 (26): 10432–10436. Bibcode:2011PNAS..10810432F. doi:10.1073/pnas.1106638108. PMC 3127884. PMID 21646521.
- ^ Augusti J, Lordkipanidze D (June 2011). "How "African" was the early human dispersal out of Africa?". Quaternary Science Reviews. 30 (11–12): 1338–1342. Bibcode:2011QSRv...30.1338A. doi:10.1016/j.quascirev.2010.04.012.
- ^ Rightmire GP (1998). "Human Evolution in the Middle Pleistocene: The Role of Homo heidelbergensis". Evolutionary Anthropology. 6 (6): 218–227. doi:10.1002/(sici)1520-6505(1998)6:6<218::aid-evan4>3.0.co;2-6. S2CID 26701026.
- ^ Asfaw B, Gilbert WH, Beyene Y, Hart WK, Renne PR, WoldeGabriel G, et al. (March 2002). "Remains of Homo erectus from Bouri, Middle Awash, Ethiopia". Nature. 416 (6878): 317–320. Bibcode:2002Natur.416..317A. doi:10.1038/416317a. PMID 11907576. S2CID 4432263.
- ^ Husson, Laurent; Salles, Tristan; Lebatard, Anne-Elisabeth; Zerathe, Swann; Braucher, Régis; Noerwidi, Sofwan; Aribowo, Sonny; Mallard, Claire; Carcaillet, Julien; Natawidjaja, Danny H.; Bourlès, Didier; ASTER team; Aumaitre, Georges; Bourlès, Didier; Keddadouche, Karim (8 November 2022). "Javanese Homo erectus on the move in SE Asia circa 1.8 Ma". Scientific Reports. 12 (1): 19012. Bibcode:2022NatSR..1219012H. doi:10.1038/s41598-022-23206-9. ISSN 2045-2322. PMC 9643487. PMID 36347897.
- ^ Zaim Y, Ciochon RL, Polanski JM, Grine FE, Bettis EA, Rizal Y, et al. (October 2011). "New 1.5 million-year-old Homo erectus maxilla from Sangiran (Central Java, Indonesia)". Journal of Human Evolution. 61 (4): 363–376. Bibcode:2011JHumE..61..363Z. doi:10.1016/j.jhevol.2011.04.009. PMID 21783226.
- ^ Ciochon RL (June 2009). "The mystery ape of Pleistocene Asia". Nature. 459 (7249): 910–911. Bibcode:2009Natur.459..910C. doi:10.1038/459910a. PMID 19536242. S2CID 205047272.
- ^ a b Kaifu Y, Baba H, Aziz F, Indriati E, Schrenk F, Jacob T (December 2005). "Taxonomic affinities and evolutionary history of the Early Pleistocene hominids of Java: dentognathic evidence". American Journal of Physical Anthropology. 128 (4): 709–726. doi:10.1002/ajpa.10425. PMID 15761880.
- ^ Perkins S (2013). "Skull suggests three early human species were one". Nature. doi:10.1038/nature.2013.13972. S2CID 88314849.
- ^ Lordkipanidze D, Ponce de León MS, Margvelashvili A, Rak Y, Rightmire GP, Vekua A, Zollikofer CP (October 2013). "A complete skull from Dmanisi, Georgia, and the evolutionary biology of early Homo". Science. 342 (6156): 326–331. Bibcode:2013Sci...342..326L. doi:10.1126/science.1238484. PMID 24136960. S2CID 20435482.
- ^ Black R (17 October 2013). "Beautiful Skull Spurs Debate on Human History". National Geographic. Archived from the original on 6 June 2021. Retrieved 6 June 2021.
- ^ Sample I (17 October 2013). "Skull of Homo erectus throws story of human evolution into disarray". The Guardian.
- ^ Giumares SW, Merino CL (September 2015). "Dmanisi hominin fossils and the problem of multiple species in the early Homo genus" (PDF). Nexus: The Canadian Student Journal of Anthropology. 23. S2CID 73528018. Archived from the original (PDF) on 14 January 2020.
- ^ Argue D, Groves CP, Lee MS, Jungers WL (June 2017). "The affinities of Homo floresiensis based on phylogenetic analyses of cranial, dental, and postcranial characters". Journal of Human Evolution. 107: 107–133. Bibcode:2017JHumE.107..107A. doi:10.1016/j.jhevol.2017.02.006. PMID 28438318.
- ^ Lordkipanidze D (4 October 2018). "Dmanisi". In Trevathan W, Cartmill M, Dufour D, Larsen C (eds.). <SCP>D</SCP> manisi. The International Encyclopedia of Biological Anthropology. John Wiley & Sons, Inc. pp. 1–4. doi:10.1002/9781118584538.ieba0139. ISBN 9781118584422. S2CID 240090147.
- ^ Baab K (December 2015). "Defining Homo erectus". Handbook of Paleoanthropology (2 ed.). pp. 2189–2219. doi:10.1007/978-3-642-39979-4_73. ISBN 978-3-642-39978-7.
- ^ Tattersall I, Schwartz J (2001). Extinct Humans. Boulder, Colorado: Westview/Perseus. ISBN 978-0-8133-3482-0.[page needed]
- ^ Kaifu Y, Aziz F, Indriati E, Jacob T, Kurniawan I, Baba H (October 2008). "Cranial morphology of Javanese Homo erectus: new evidence for continuous evolution, specialization, and terminal extinction". Journal of Human Evolution. 55 (4): 551–580. Bibcode:2008JHumE..55..551K. doi:10.1016/j.jhevol.2008.05.002. PMID 18635247.
- ^ Vlček, Emanuel (1 March 1978). "A new discovery of Homo erectus in central Europe". Journal of Human Evolution. 7 (3): 239–251. Bibcode:1978JHumE...7..239V. doi:10.1016/S0047-2484(78)80115-8. ISSN 0047-2484.
- ^ Arsuaga, Juan-Luis; Martínez, Ignacio; Gracia, Ana; Carretero, José-Miguel; Carbonell, Eudald (1993). "Three new human skulls from the Sima de los Huesos Middle Pleistocene site in Sierra de Atapuerca, Spain". Nature. 362 (6420): 534–537. Bibcode:1993Natur.362..534A. doi:10.1038/362534a0. ISSN 1476-4687. PMID 8464493. S2CID 4321154.
- ^ a b c Coon, Carleton S. (1966). "Review of The Nomenclature of the Hominidae, including a Definitive List of Hominid Taxa". Human Biology. 38 (3): 344–347. ISSN 0018-7143. JSTOR 41449277.
- ^ Singer, Ronald (1958). "232. The Boskop 'Race' Problem". Man. 58: 173–178. doi:10.2307/2795854. ISSN 0025-1496. JSTOR 2795854.
- ^ a b 李学勤 (2007). 20世纪中国学术大典: 考古学, 博物馆学 (in Chinese). 福建教育出版社. ISBN 978-7-5334-3641-4.
- ^ E, Dubois (1891). "Palaeontologische onderzoekingen op Java". Verslag van het Mijnwezen, 3e/4e Kwartaal: 12–15.
- ^ Tuttle, Russell H. (12 May 2011). Paleoanthropology: Morphology and Paleoecology. Walter de Gruyter. ISBN 978-3-11-081069-1.
- ^ a b c d Antón, Susan C.; Middleton, Emily R. (1 June 2023). "Making meaning from fragmentary fossils: Early Homo in the Early to early Middle Pleistocene". Journal of Human Evolution. 179: 103307. Bibcode:2023JHumE.17903307A. doi:10.1016/j.jhevol.2022.103307. ISSN 0047-2484. PMID 37030994.
- ^ Schwartz, Jeffrey H.; Tattersall, Ian; Chi, Zhang (25 April 2014). "Comment on "A Complete Skull from Dmanisi, Georgia, and the Evolutionary Biology of Early Homo "". Science. 344 (6182): 360. Bibcode:2014Sci...344..360S. doi:10.1126/science.1250056. ISSN 0036-8075. PMID 24763572. S2CID 36578190.
- ^ Gabounia, Léo; de Lumley, Marie-Antoinette; Vekua, Abesalom; Lordkipanidze, David; de Lumley, Henry (1 September 2002). "Découverte d'un nouvel hominidé à Dmanissi (Transcaucasie, Géorgie)". Comptes Rendus Palevol (in French). 1 (4): 243–253. Bibcode:2002CRPal...1..243G. doi:10.1016/S1631-0683(02)00032-5. ISSN 1631-0683.
- ^ Rightmire, G. Philip; Lordkipanidze, David; Vekua, Abesalom (1 February 2006). "Anatomical descriptions, comparative studies and evolutionary significance of the hominin skulls from Dmanisi, Republic of Georgia". Journal of Human Evolution. 50 (2): 115–141. Bibcode:2006JHumE..50..115R. doi:10.1016/j.jhevol.2005.07.009. ISSN 0047-2484. PMID 16271745.
- ^ Rukang, Wu (15 June 1992). "On the classification of subspecies of Homo". Acta Anthropologica Sinica. 11 (2): 109. ISSN 1000-3193.
- ^ Tobias, Phillip V. (1991). "The species Homo habilis: example of a premature discovery". Annales Zoologici Fennici. 28 (3/4): 371–380. ISSN 0003-455X. JSTOR 23735461.
- ^ Mounier, A.; Caparros, M. (1 October 2015). "Le statut phylogénétique d'Homo heidelbergensis – étude cladistique des homininés du Pléistocène moyen". BMSAP (in French). 27 (3): 110–134. doi:10.1007/s13219-015-0127-4. ISSN 1777-5469. S2CID 17449909.
- ^ W, Huang (1982). "Preliminary study on the fossil hominid skull and fauna of Hexian, Anhui". Vertebrata PalAsiatica. 20: plate 1.
- ^ Naddeo, Michelangelo (2023). "Az ősi magyar jelképrendszer keresése" (PDF). Acta Historica Hungaricus. 38.
- ^ Woo, J.-K. (1964). "The Hominid Skull of Lantian, Shenshi" (PDF). Vertebrata PalAsiatica. 10 (1).
- ^ Groves, Colin P. (1 December 1999). "Nomenclature of African Plio-Pleistocene hominins". Journal of Human Evolution. 37 (6): 869–872. Bibcode:1999JHumE..37..869G. doi:10.1006/jhev.1999.0366. ISSN 0047-2484. PMID 10600324.
- ^ "Homo erectus mapaensis Kurth, 1965". Paleo Core. 2023. Retrieved 7 August 2023.
- ^ Sonakia, Arun; Kennedy, Kenneth A. R. (1985). "Skull Cap of an Early Man from the Narmada Valley Alluvium (Pleistocene) of Central India". American Anthropologist. 87 (3): 612–616. doi:10.1525/aa.1985.87.3.02a00060. ISSN 0002-7294. JSTOR 678879.
- ^ Laitman, Jeffrey T.; Tattersall, Ian (1 April 2001). "Homo erectus newyorkensis: An Indonesian fossil rediscovered in Manhattan sheds light on the middle phase of human evolution". The Anatomical Record. 262 (4): 341–343. doi:10.1002/ar.1042. ISSN 0003-276X. PMID 11275967. S2CID 35310160.
- ^ SARTONO R (1976). The Javanese Pleistocene Hominids: A Re-Appraisal.
- ^ Vlček, Emanuel; Mania, Dietrich (1977). "Ein Neuer Fund Von Homo Erectus in Europa: Bilzingsleben (ddr)". Anthropologie (1962-). 15 (2/3): 159–169. ISSN 0323-1119. JSTOR 26294532.
- ^ D, Black (1927). "On a lower molar hominid tooth from the Chou Kou Tien deposit". Palaeont. Sinica, Ser. D. 7: 1–29.
- ^ Dean, David; Hublin, Jean-Jacques; Holloway, Ralph; Ziegler, Reinhard (1998). "On the phylogenetic position of the pre-Neandertal specimen from Reilingen, Germany". Journal of Human Evolution. 34 (5): 485–508. Bibcode:1998JHumE..34..485D. doi:10.1006/jhev.1998.0214. PMID 9614635.
- ^ Oppenoorth, W.F.F. (1932). "Homo (Javanthropus) soloensis. Ein plistocene mensch van Java". Wetenschappelijke Mededeelingen. 20: 49–75.
- ^ Guipert, Gaspard (30 November 2005). Reconstitution et position phylétique des restes crâniens de l'Homme de Tautavel (Arago 21-47) et de Biache-Saint-Vaast 2. Apports de l'imagerie et de l'analyse tridimensionnelle (phdthesis thesis) (in French). Université Paul Cézanne - Aix-Marseille III.
- ^ Sartono, S. (1980). "On the Javanese Pleistocene hominids: A reappraisal". Abstracts of the IUSPP Nice.
- ^ Dubois, E. (1 January 1921). "The proto-Australian fossil man of Wadjak, Java". Koninklijke Nederlandse Akademie van Wetenschappen Proceedings Series B Physical Sciences. 23: 1013–1051. Bibcode:1921KNAB...23.1013D.
- ^ Pramujiono, Agung. "BERBAGAI PANDANGAN ASAL BANGSA DAN BAHASA INDONESIA: DARI KAJIAN LINGUSITIK HISTORIS KOMPARATIF SAMPAI ARKEOLINGUISTIK DAN PALEOLINGUISTIK". Academia.edu.
- ^ Wei, Guangbiao; Huang, Wanbo; Chen, Shaokun; He, Cunding; Pang, Libo; Wu, Yan (15 December 2014). "Paleolithic culture of Longgupo and its creators". Quaternary International. Multidisciplinary Perspectives on the Gigantopithecus Fauna and Quaternary Biostratigraphy in East Asia. 354: 154–161. Bibcode:2014QuInt.354..154W. doi:10.1016/j.quaint.2014.04.003. ISSN 1040-6182.
- ^ Zanolli, Clément; Kullmer, Ottmar; Kelley, Jay; Bacon, Anne-Marie; Demeter, Fabrice; Dumoncel, Jean; Fiorenza, Luca; Grine, Frederick E.; Hublin, Jean-Jacques; Nguyen, Anh Tuan; Nguyen, Thi Mai Huong; Pan, Lei; Schillinger, Burkhard; Schrenk, Friedemann; Skinner, Matthew M. (May 2019). "Evidence for increased hominid diversity in the Early to Middle Pleistocene of Indonesia". Nature Ecology & Evolution. 3 (5): 755–764. Bibcode:2019NatEE...3..755Z. doi:10.1038/s41559-019-0860-z. ISSN 2397-334X. PMID 30962558. S2CID 102353734.
- ^ Chengzhi, Hu (1973). "Yunnan Yuanmou faxian de yuanren yachi huashi". Dizhi Xuebao. 1: 65–71.
- ^ There was long-standing uncertainty whether H. floresiensis should be considered close to H. erectus, close to H. sapiens, or an altogether separate species. In 2017, it was suggested on morphological grounds that H. floresiensis is a sister species to either H. habilis or to a minimally habilis-erectus-ergaster-sapiens clade, and its line much more ancient than Homo erectus itself. Argue D, Groves CP, Lee MS, Jungers WL (June 2017). "The affinities of Homo floresiensis based on phylogenetic analyses of cranial, dental, and postcranial characters". Journal of Human Evolution. 107: 107–133. Bibcode:2017JHumE.107..107A. doi:10.1016/j.jhevol.2017.02.006. PMID 28438318.
- ^ Kennedy KA, Sonakia A, Chiment J, Verma KK (December 1991). "Is the Narmada hominid an Indian Homo erectus?". American Journal of Physical Anthropology. 86 (4): 475–496. doi:10.1002/ajpa.1330860404. PMID 1776655.
- ^ Krantz, G.S. (1975). "An explanation for the diastema of Javan erectus Skull IV". In: Paleoanthropology, Morphology and Paleoecology. La Hague: Mouton, 361–372.
- ^ Zanolli C, Kullmer O, Kelley J, Bacon AM, Demeter F, Dumoncel J, et al. (May 2019). "Evidence for increased hominid diversity in the Early to Middle Pleistocene of Indonesia" (PDF). Nature Ecology & Evolution. 3 (5): 755–764. Bibcode:2019NatEE...3..755Z. doi:10.1038/s41559-019-0860-z. PMID 30962558. S2CID 102353734.
- ^ Baba H, Aziz F, Kaifu Y, Suwa G, Kono RT, Jacob T (February 2003). "Homo erectus calvarium from the Pleistocene of Java". Science. 299 (5611): 1384–1388. doi:10.1126/science.1081676. PMID 12610302. S2CID 20437090.
- ^ a b Balzeau A (2006). "Are thickened cranial bones and equal participation of the three structural bone layers autapomorphic traits of Homo erectus?". Bulletins et mémoires de la Société d'Anthropologie de Paris. 18 (3–4): 145–163. doi:10.4000/bmsap.1528.
- ^ Copes LE, Kimbel WH (January 2016). "Cranial vault thickness in primates: Homo erectus does not have uniquely thick vault bones". Journal of Human Evolution. 90: 120–134. Bibcode:2016JHumE..90..120C. doi:10.1016/j.jhevol.2015.08.008. PMID 26767964.
- ^ Franciscus RG, Trinkaus E (April 1988). "Nasal morphology and the emergence of Homo erectus". American Journal of Physical Anthropology. 75 (4): 517–527. doi:10.1002/ajpa.1330750409. PMID 3133950.
- ^ Jacobs LF (February 2019). "The navigational nose: a new hypothesis for the function of the human external pyramid". The Journal of Experimental Biology. 222 (Pt Suppl 1): jeb186924. doi:10.1242/jeb.186924. PMID 30728230.
- ^ a b Antón SC, Taboada HG, Middleton ER, Rainwater CW, Taylor AB, Turner TR, et al. (July 2016). "Morphological variation in Homo erectus and the origins of developmental plasticity". Philosophical Transactions of the Royal Society of London. Series B, Biological Sciences. 371 (1698): 20150236. doi:10.1098/rstb.2015.0236. PMC 4920293. PMID 27298467.
- ^ Clark, Gary; Henneberg, Maciej (2022). "Interpopulational variation in human brain size: Implications for hominin cognitive phylogeny". Anthropological Review. 84 (4): 405–429. doi:10.2478/anre-2021-0029.
- ^ a b c d e Ungar PS, Grine FE (2006). "Diet in Early Homo: A Review of the Evidence and a New Model of Adaptive Versatility". Annual Review of Anthropology. 35: 208–228. doi:10.1146/annurev.anthro.35.081705.123153.
- ^ Alan Walker, Richard Leakey (1993). The Nariokotome Homo erectus skeleton. Harvard University Press. p. 412. ISBN 9780674600751. Retrieved 2 October 2022.
- ^ Migliano AB, Guillon M (2012). "The Effects of Mortality, Subsistence, and Ecology on Human Adult Height and Implications for Homo Evolution". Current Anthropology. 53 (S6): 359–368. doi:10.1086/667694. S2CID 84442763.
- ^ a b Antón, Susan C.; Taboada, Hannah G.; Middleton, Emily R.; Rainwater, Christopher W.; Taylor, Andrea B.; Turner, Trudy R.; Turnquist, Jean E.; Weinstein, Karen J.; Williams, Scott A. (5 July 2016). "Morphological variation in Homo erectus and the origins of developmental plasticity". Philosophical Transactions of the Royal Society B: Biological Sciences. 371 (1698): 20150236. doi:10.1098/rstb.2015.0236. ISSN 0962-8436. PMC 4920293. PMID 27298467.
- ^ Antón, Susan C.; Potts, Richard; Aiello, Leslie C. (4 July 2014). "Evolution of early Homo: An integrated biological perspective". Science. 345 (6192): 1236828. doi:10.1126/science.1236828. ISSN 0036-8075. PMID 24994657. S2CID 30188239.
- ^ Antón, Susan C.; Taboada, Hannah G.; Middleton, Emily R.; Rainwater, Christopher W.; Taylor, Andrea B.; Turner, Trudy R.; Turnquist, Jean E.; Weinstein, Karen J.; Williams, Scott A. (5 July 2016). "Morphological variation in Homo erectus and the origins of developmental plasticity". Philosophical Transactions of the Royal Society B: Biological Sciences. 371 (1698): 20150236. doi:10.1098/rstb.2015.0236. ISSN 0962-8436. PMC 4920293. PMID 27298467.
- ^ a b c Simpson SW, Quade J, Levin NE, Butler R, Dupont-Nivet G, Everett M, Semaw S (November 2008). "A female Homo erectus pelvis from Gona, Ethiopia". Science. 322 (5904): 1089–1092. Bibcode:2008Sci...322.1089S. doi:10.1126/science.1163592. PMID 19008443. S2CID 22191315.
- ^ a b Leakey, Mary D (1979). Olduvai Gorge: my search for early man. London: Collins. ISBN 9780002116138. OCLC 647137093.
- ^ a b Plavcan JM (2012). "Body Size, Size Variation, and Sexual Size Dimorphism in Early Homo". Current Anthropology. 53 (S6): 309–423. doi:10.1086/667605. S2CID 84095311.
- ^ "Rightmire GP. The Evolution of Homo Erectus: Comparative Anatomical Studies of an Extinct Human Species. Cambridge University Press; 1990". search.library.ucr.edu. Retrieved 5 May 2022.
- ^ Ruff C (March 2008). "Femoral/humeral strength in early African Homo erectus". Journal of Human Evolution. 54 (3): 383–390. Bibcode:2008JHumE..54..383R. doi:10.1016/j.jhevol.2007.09.001. PMID 17977577.
- ^ a b Hatala KG, Roach NT, Ostrofsky KR, Wunderlich RE, Dingwall HL, Villmoare BA, et al. (July 2016). "Footprints reveal direct evidence of group behavior and locomotion in Homo erectus". Scientific Reports. 6 (28766): 28766. Bibcode:2016NatSR...628766H. doi:10.1038/srep28766. PMC 4941528. PMID 27403790.
- ^ Roach, & Richmond. (2015). "Clavicle length, throwing performance and the reconstruction of the Homo erectus shoulder". Journal of Human Evolution, 80(C), 107–113.
- ^ Haeusler M, Schiess R, Boeni T (November 2011). "New vertebral and rib material point to modern bauplan of the Nariokotome Homo erectus skeleton" (PDF). Journal of Human Evolution. 61 (5): 575–582. Bibcode:2011JHumE..61..575H. doi:10.1016/j.jhevol.2011.07.004. PMID 21868059.
- ^ Rogers AR, Iltis D, Wooding S (2004). "Genetic Variation at the MC1R Locus and the Time since Loss of Human Body Hair". Current Anthropology. 45 (1): 105–108. doi:10.1086/381006. S2CID 224795768.
- ^ a b Gilligan I (2010). "The Prehistoric Development of Clothing: Archaeological Implications of a Thermal Model". Journal of Archaeological Method and Theory. 15: 15–80. doi:10.1007/s10816-009-9076-x. S2CID 143004288.
- ^ Dávid-Barrett T, Dunbar RI (May 2016). "Bipedality and hair loss in human evolution revisited: The impact of altitude and activity scheduling". Journal of Human Evolution. 94: 72–82. Bibcode:2016JHumE..94...72D. doi:10.1016/j.jhevol.2016.02.006. PMC 4874949. PMID 27178459.
- ^ Jablonski NG (March 2012). "Human skin pigmentation as an example of adaptive evolution". Proceedings of the American Philosophical Society. 156 (1): 45–57. JSTOR 23558077. PMID 23035389.
- ^ a b Kappelman J, Alçiçek MC, Kazanci N, Schultz M, Ozkul M, Sen S (January 2008). "First Homo erectus from Turkey and implications for migrations into temperate Eurasia". American Journal of Physical Anthropology. 135 (1): 110–116. doi:10.1002/ajpa.20739. PMID 18067194.
- ^ Best A, Kamilar JM (April 2018). "The evolution of eccrine sweat glands in human and nonhuman primates". Journal of Human Evolution. 117: 33–43. Bibcode:2018JHumE.117...33B. doi:10.1016/j.jhevol.2017.12.003. PMID 29544622. S2CID 3921318.
- ^ Pagel M, Bodmer W (2004). "The Evolution of Human Hairlessness: Cultural Adaptations and the Ectoparasite Hypothesis". Evolutionary Theory and Processes: Modern Horizons. Springer, Dordrecht. pp. 329–335. doi:10.1007/978-94-017-0443-4_17. ISBN 978-94-017-0443-4.
- ^ Gile J (2010). "Naked Love: The Evolution of Human Hairlessness". Biological Theory. 5 (4): 326–336. doi:10.1162/BIOT_a_00062. S2CID 84164968.
- ^ a b c Coqueugniot H, Hublin JJ, Veillon F, Houët F, Jacob T (September 2004). "Early brain growth in Homo erectus and implications for cognitive ability". Nature. 431 (7006): 299–302. Bibcode:2004Natur.431..299C. doi:10.1038/nature02852. PMID 15372030. S2CID 4428043.
- ^ Caspari R, Lee SH (July 2004). "Older age becomes common late in human evolution". Proceedings of the National Academy of Sciences of the United States of America. 101 (30): 10895–10900. doi:10.1073/pnas.0402857101. PMC 503716. PMID 15252198.
- ^ Steudel-Numbers KL (November 2006). "Energetics in Homo erectus and other early hominins: the consequences of increased lower-limb length". Journal of Human Evolution. 51 (5): 445–453. Bibcode:2006JHumE..51..445S. doi:10.1016/j.jhevol.2006.05.001. PMID 16780923.
- ^ a b c d e Ben-Dor M, Gopher A, Hershkovitz I, Barkai R (2011). "Man the fat hunter: the demise of Homo erectus and the emergence of a new hominin lineage in the Middle Pleistocene (ca. 400 kyr) Levant". PLOS ONE. 6 (12): e28689. Bibcode:2011PLoSO...628689B. doi:10.1371/journal.pone.0028689. PMC 3235142. PMID 22174868.
- ^ a b Boaz N, Ciochon R (2004). "Headstrong Hominids". Natural History. 113 (1): 28–34.
- ^ a b Kennedy GE (1985). "Bone thickness in Homo erectus". Journal of Human Evolution. 14 (8): 699–708. Bibcode:1985JHumE..14..699K. doi:10.1016/S0047-2484(85)80052-X.
- ^ Russell MD, Brown T, Garn SM, Giris F, Turkel S, İşcan MY, et al. (1985). "The Supraorbital Torus: 'A Most Remarkable Peculiarity'". Current Anthropology. 26 (3): 337–350. doi:10.1086/203279. S2CID 146857927.
- ^ a b Willems EP, van Schaik CP (August 2017). "The social organization of Homo ergaster: Inferences from anti-predator responses in extant primates". Journal of Human Evolution. 109: 11–21. Bibcode:2017JHumE.109...11W. doi:10.1016/j.jhevol.2017.05.003. PMID 28688456.
- ^ a b Carotenuto F, Tsikaridze N, Rook L, Lordkipanidze D, Longo L, Condemi S, Raia P (June 2016). "Venturing out safely: The biogeography of Homo erectus dispersal out of Africa". Journal of Human Evolution. 95: 1–12. Bibcode:2016JHumE..95....1C. doi:10.1016/j.jhevol.2016.02.005. hdl:10356/82274. PMID 27260171.
- ^ Plavcan JM (2012). "Implications of Male and Female Contributions to Sexual Size Dimorphism for Inferring Behavior in the Hominin Fossil Record". International Journal of Primatology. 33 (6): 1364–1381. doi:10.1007/s10764-012-9642-z. S2CID 17850676.
- ^ Melamed Y, Kislev ME, Geffen E, Lev-Yadun S, Goren-Inbar N (December 2016). "The plant component of an Acheulian diet at Gesher Benot Ya'aqov, Israel". Proceedings of the National Academy of Sciences of the United States of America. 113 (51): 14674–14679. Bibcode:2016PNAS..11314674M. doi:10.1073/pnas.1607872113. PMC 5187744. PMID 27930293.
- ^ Steele TE (June 2010). "A unique hominin menu dated to 1.95 million years ago". Proceedings of the National Academy of Sciences of the United States of America. 107 (24): 10771–10772. Bibcode:2010PNAS..10710771S. doi:10.1073/pnas.1005992107. PMC 2890732. PMID 20534542.
- ^ Patterson, D. B.; Braun, D. R.; Behrensmeyer, A. K.; Merritt, S.; Zliobaite, I.; Reeves, J. S.; Wood, B. A.; Fortelius, M.; Bobe, R. (1 September 2017). "Ecosystem evolution and hominin paleobiology at East Turkana, northern Kenya between 2.0 and 1.4 Ma". Palaeogeography, Palaeoclimatology, Palaeoecology. 481: 1–13. Bibcode:2017PPP...481....1P. doi:10.1016/j.palaeo.2017.05.001.
- ^ Joordens JC, Wesselingh FP, de Vos J, Vonhof HB, Kroon D (December 2009). "Relevance of aquatic environments for hominins: a case study from Trinil (Java, Indonesia)". Journal of Human Evolution. 57 (6): 656–671. Bibcode:2009JHumE..57..656J. doi:10.1016/j.jhevol.2009.06.003. PMID 19683789.
- ^ a b c d e f Gowlett JA (June 2016). "The discovery of fire by humans: a long and convoluted process". Philosophical Transactions of the Royal Society of London. Series B, Biological Sciences. 371 (1696): 20150164. doi:10.1098/rstb.2015.0164. PMC 4874402. PMID 27216521.
- ^ a b c d e Gowlett JA, Wrangham RW (2013). "Earliest fire in Africa: Towards the convergence of archaeological evidence and the cooking hypothesis". Azania: Archaeological Research in Africa. 48 (1): 5–30. doi:10.1080/0067270X.2012.756754. S2CID 163033909.
- ^ Rhodin A, Thomson S, Georgalis G, Karl HV, Danilov I, Takahashi A, de la Fuente M, Bourque J, Delfino M, Bour R, Iverson J, Shaffer B, Van Dijk PP (2015). "Turtles and Tortoises of the World During the Rise and Global Spread of Humanity: First Checklist and Review of Extinct Pleistocene and Holocene Chelonians". In Rhodin A, Pritchard P, van Dijk PP, Saumure R, Buhlmann K, Iverson J, Mittermeier R (eds.). Conservation Biology of Freshwater Turtles and Tortoises. Chelonian Research Monographs. Vol. 5 (First ed.). Chelonian Research Foundation. p. 15. doi:10.3854/crm.5.000e.fossil.checklist.v1.2015. ISBN 978-0-9653540-9-7.
- ^ Beck RB, Black L, Krieger LS, Naylor PC, Shabaka DI (1999). World History: Patterns of Interaction. Evanston, IL: McDougal Littell. ISBN 978-0-395-87274-1.[page needed]
- ^ Richards MP (December 2002). "A brief review of the archaeological evidence for Palaeolithic and Neolithic subsistence". European Journal of Clinical Nutrition. 56 (12): 1270–1278. doi:10.1038/sj.ejcn.1601646. PMID 12494313.
- ^ de la Torre I (July 2016). "The origins of the Acheulean: past and present perspectives on a major transition in human evolution". Philosophical Transactions of the Royal Society of London. Series B, Biological Sciences. 371 (1698): 20150245. doi:10.1098/rstb.2015.0245. PMC 4920301. PMID 27298475.
- ^ a b Lepre CJ, Roche H, Kent DV, Harmand S, Quinn RL, Brugal JP, et al. (August 2011). "An earlier origin for the Acheulian". Nature. 477 (7362): 82–85. Bibcode:2011Natur.477...82L. doi:10.1038/nature10372. PMID 21886161. S2CID 4419567.
- ^ a b c Nowell A, Chang ML (2009). "The Case Against Sexual Selection as an Explanation of Handaxe Morphology" (PDF). PaleoAnthropology: 77–88.
- ^ Wynn T (1979). "The Intelligence of Later Acheulean Hominids". Man. 14 (3): 371–391. doi:10.2307/2801865. JSTOR 2801865.
- ^ Choi K, Driwantoro D (2007). "Shell tool use by early members of Homo erectus in Sangiran, central Java, Indonesia: cut mark evidence". Journal of Archaeological Science. 34 (1): 48–58. Bibcode:2007JArSc..34...48C. doi:10.1016/j.jas.2006.03.013.
- ^ a b Joordens JC, d'Errico F, Wesselingh FP, Munro S, de Vos J, Wallinga J, et al. (February 2015). "Homo erectus at Trinil on Java used shells for tool production and engraving". Nature. 518 (7538): 228–231. Bibcode:2015Natur.518..228J. doi:10.1038/nature13962. PMID 25470048. S2CID 4461751.
- ^ Turner J (1996). History and Science of Knots. World Scientific. pp. 6–8. ISBN 9789810224691.
- ^ Hlubik S, Berna F, Feibel C, Braun D (2017). "Researching the Nature of Fire at 1.5 Mya on the Site of FxJj20 AB, Koobi Fora, Kenya, Using High-Resolution Spatial Analysis and FTIR Spectrometry". Current Anthropology. 58: S243–S257. doi:10.1086/692530. S2CID 148948219.
- ^ a b c d Roebroeks W, Villa P (March 2011). "On the earliest evidence for habitual use of fire in Europe". Proceedings of the National Academy of Sciences of the United States of America. 108 (13): 5209–5214. Bibcode:2011PNAS..108.5209R. doi:10.1073/pnas.1018116108. PMC 3069174. PMID 21402905.
- ^ Beaumont PB (2011). "The Edge: More on Fire-Making by about 1.7 Million Years Ago at Wonderwerk Cave in South Africa". Current Anthropology. 52 (4): 585–595. doi:10.1086/660919. S2CID 144176681.
- ^ a b Sandgathe D (2017). "Identifying and Describing Pattern and Process in the Evolution of Hominin Use of Fire". Current Anthropology. 58: S360–S370. doi:10.1086/691459. hdl:11858/00-001M-0000-002C-0141-3. S2CID 165025762.
- ^ Antón SC (2003). "Natural history of Homo erectus". American Journal of Physical Anthropology. 122 (S37): 126–170. doi:10.1002/ajpa.10399. PMID 14666536.
- ^ Zhong M, Shi C, Gao X, Wu X, Chen F, Zhang S, Zhang X, Olsen JW (2013). "On the possible use of fire by Homo erectus at Zhoukoudian, China". Chinese Science Bulletin. 59 (3): 335–343. doi:10.1007/s11434-013-0061-0. S2CID 93590269.
- ^ Musée de Préhistoire Terra Amata. "Le site acheuléen de Terra Amata" [The Acheulean site of Terra Amata]. Musée de Préhistoire Terra Amata (in French). Retrieved 10 June 2022.
- ^ Leakey MD (1971). Olduvai Gorge: Volume 3, Excavations in Beds I and II, 1960-1963. Cambridge University Press. p. 24. ISBN 9780521077231.
- ^ Ingold T (2000). "Building, dwelling, living: how animals and people make themselves at home in the world". The Perception of the Environment: Essays on Livelihood, Dwelling and Skill. Psychology Press. p. 184. ISBN 9780415228329.
- ^ Ullman M, Hovers E, Goren-Inbar N, Frumkin A (2013). "Levantine cave dwellers: geographic and environmental aspects of early humans use of caves, case study from Wadi Amud, northern Israel". International Congress of Speleology. 1.
- ^ Toups MA, Kitchen A, Light JE, Reed DL (January 2011). "Origin of clothing lice indicates early clothing use by anatomically modern humans in Africa". Molecular Biology and Evolution. 28 (1): 29–32. doi:10.1093/molbev/msq234. PMC 3002236. PMID 20823373.
- ^ "New evidence in search for the mysterious Denisovans". ScienceDaily. 23 March 2021. Retrieved 30 March 2021.
- ^ a b Bednarik RG (1999). "Pleistocene seafaring in the Mediterranean". Anthropologie. 37 (3): 275–282. JSTOR 26294895.
- ^ Bednarik RG (1998). "An experiment in Pleistocene seafaring" (PDF). The International Journal of Nautical Archaeology. 27 (2): 139–149. Bibcode:1998IJNAr..27..139B. doi:10.1111/j.1095-9270.1998.tb00797.x.
- ^ Détroit F, Mijares AS, Corny J, Daver G, Zanolli C, Dizon E, et al. (April 2019). "A new species of Homo from the Late Pleistocene of the Philippines" (PDF). Nature. 568 (7751): 181–186. Bibcode:2019Natur.568..181D. doi:10.1038/s41586-019-1067-9. PMID 30971845. S2CID 106411053.
- ^ Spikins P, Needham A, Wright B, Dytham C, Gatta M, Hitchens G (2019). "Living to fight another day: The ecological and evolutionary significance of Neanderthal healthcare". Quaternary Science Reviews. 217: 98–118. Bibcode:2019QSRv..217...98S. doi:10.1016/j.quascirev.2018.08.011.
- ^ Lordkipanidze D, Vekua A, Ferring R, Rightmire GP, Agusti J, Kiladze G, et al. (April 2005). "Anthropology: the earliest toothless hominin skull". Nature. 434 (7034): 717–718. Bibcode:2005Natur.434..717L. doi:10.1038/434717b. PMID 15815618. S2CID 52800194.
- ^ Haeusler M, Schiess R, Boeni T (February 2013). "Evidence for juvenile disc herniation in a homo erectus boy skeleton" (PDF). Spine. 38 (3): E123–E128. doi:10.1097/BRS.0b013e31827cd245. PMID 23154836. S2CID 11534863.
- ^ Soriano M (January 1970). "The fluoric origin of the bone lesion in the Pithecanthropus erectus femur". American Journal of Physical Anthropology. 32 (1): 49–57. doi:10.1002/ajpa.1330320107. PMID 4984453.
- ^ Henshilwood CS, d'Errico F, Watts I (July 2009). "Engraved ochres from the Middle Stone Age levels at Blombos Cave, South Africa". Journal of Human Evolution. 57 (1): 27–47. Bibcode:2009JHumE..57...27H. doi:10.1016/j.jhevol.2009.01.005. PMID 19487016.
- ^ d'Errico F, Moreno RG, Rifkin RF (2012). "Technological, elemental and colorimetric analysis of an engraved ochre fragment from the Middle Stone Age levels of Klasies River Cave 1, South Africa". J. Archaeol. Sci. 39 (4): 942–952. Bibcode:2012JArSc..39..942D. doi:10.1016/j.jas.2011.10.032.
- ^ Callaway E (2014). "Homo erectus made world's oldest doodle 500,000 years ago". Nature News. doi:10.1038/nature.2014.16477. S2CID 164153158.
- ^ a b c d e Dickson DB (1992). The Dawn of Belief: Religion in the Upper Paleolithic of Southwestern Europe. University of Arizona Press. pp. 40–46. ISBN 978-0-8165-1336-9.
- ^ a b Morriss-Kay GM (February 2010). "The evolution of human artistic creativity". Journal of Anatomy. 216 (2): 158–176. doi:10.1111/j.1469-7580.2009.01160.x. PMC 2815939. PMID 19900185.
- ^ d'Errico F, Nowell A (2000). "A New Look at the Berekhat Ram Figurine: Implications for the Origins of Symbolism". Cambridge Archaeological Journal. 10 (1): 123–167. doi:10.1017/S0959774300000056. S2CID 163138037.
- ^ a b Watts I (2014). "The red thread: pigment use and the evolution of collective ritual". The Social Origins of Language. Oxford University Press. pp. 222–223. ISBN 978-0-19-966533-4.
- ^ de Lumley H, Boone Y (1976). "Les structures d'habitat au Paléolithique moyen" [Housing structures from the lower Paleolithic]. In de Lumley H, Guilaine J (eds.). La Préhistoire française: Les civilisations paléolithiques et mésolithiques de la France [French prehistory: the Paleolithic and Mesolithic civilizations of France]. Éditions du Centre national de la recherche scientifique. ISBN 978-2-222-01968-8.
- ^ Wreschner EE, Bolton R, Butzer KW, Delporte H, Häusler A, Heinrich A, et al. (1980). "Red Ochre and Human Evolution: A Case for Discussion" (PDF). Current Anthropology. 21 (5): 632–633. doi:10.1086/202541. JSTOR 2741829. S2CID 88099778.
- ^ Weidenreich F (1935). "The Sinanthropus Population of Choukoutien (Locality 1) with a Preliminary Report on New Discoveries". Bulletin of the Geological Society of China. 14 (4): 427–468. doi:10.1111/j.1755-6724.1935.mp14004001.x.
- ^ Binford LR, Ho CK (1985). "Taphonomy at a Distance: Zhoukoudian, 'The Cave Home of Beijing Man'?". Current Anthropology. 26 (4): 413–442. doi:10.1086/203303. JSTOR 2742759. S2CID 147164100.
- ^ Kohn M, Mithen S (1999). "Handaxes: products of sexual selection?". Antiquity. 73 (281): 518–526. doi:10.1017/S0003598X00065078. S2CID 162903453.
- ^ Latimer B, Ohman J (2001). "Axial dysplasia in Homo erectus". Journal of Human Evolution. 40.
- ^ Meyer M, Lordkipanidze D, Vekua A (2006). Language and empathy in Homo erectus: Behaviors suggested by a modern spinal cord from Dmanisi, but not Nariokotome. Annual meeting of the Paleoanthroplogy Society. San Juan, Puerto Rico.
- ^ Schiess R, Haeusler M (March 2013). "No skeletal dysplasia in the Nariokotome boy KNM-WT 15000 (Homo erectus)--a reassessment of congenital pathologies of the vertebral column". American Journal of Physical Anthropology. 150 (3): 365–374. doi:10.1002/ajpa.22211. PMID 23283736.
- ^ Schiess R, Boeni T, Rühli F, Haeusler M (February 2014). "Revisiting scoliosis in the KNM-WT 15000 Homo erectus skeleton" (PDF). Journal of Human Evolution. 67 (48–59): 48–59. Bibcode:2014JHumE..67...48S. doi:10.1016/j.jhevol.2013.12.009. PMID 24491377.
- ^ Luef EM (2018). "Tracing the human brain's classical language areas in extant and extinct hominids". The talking species: Perspectives on the evolutionary, neuronal and cultural foundations of language. Uni-Press Graz. ISBN 978-3-902666-52-9.
- ^ Capasso L, Michetti E, D'Anastasio R (December 2008). "A Homo erectus hyoid bone: possible implications for the origin of the human capability for speech". Collegium Antropologicum. 32 (4): 1007–1011. PMID 19149203.
- ^ Hillert DG (2015). "On the Evolving Biology of Language". Frontiers in Psychology. 6: 1796. doi:10.3389/fpsyg.2015.01796. PMC 4656830. PMID 26635694.
- ^ Martínez I, Arsuaga JL, Quam R, Carretero JM, Gracia A, Rodríguez L (January 2008). "Human hyoid bones from the middle Pleistocene site of the Sima de los Huesos (Sierra de Atapuerca, Spain)" (PDF). Journal of Human Evolution. 54 (1): 118–124. Bibcode:2008JHumE..54..118M. doi:10.1016/j.jhevol.2007.07.006. PMID 17804038.
- ^ Martínez I, Rosa M, Arsuaga JL, Jarabo P, Quam R, Lorenzo C, et al. (July 2004). "Auditory capacities in Middle Pleistocene humans from the Sierra de Atapuerca in Spain". Proceedings of the National Academy of Sciences of the United States of America. 101 (27): 9976–9981. Bibcode:2004PNAS..101.9976M. doi:10.1073/pnas.0403595101. PMC 454200. PMID 15213327.
- ^ Padmanaban D (6 November 2020). "Climate Change May Have Been a Major Driver of Ancient Hominin Extinctions". SAPIENS. Retrieved 9 November 2020.
- ^ "Climate change likely drove early human species to extinction, modeling study suggests". phys.org. Retrieved 9 November 2020.
- ^ Raia P, Mondanaro A, Melchionna M, Di Febbraro M, Diniz-Filho JA, Rangel TF, et al. (23 October 2020). "Past Extinctions of Homo Species Coincided with Increased Vulnerability to Climatic Change". One Earth. 3 (4): 480–490. Bibcode:2020OEart...3..480R. doi:10.1016/j.oneear.2020.09.007. hdl:2158/1211341. ISSN 2590-3330.
- ^ a b c Zanolli, Clément, et al. "Inner Tooth Morphology of Homo Erectus from Zhoukoudian. New Evidence from an Old Collection Housed at Uppsala University, Sweden." Journal of Human Evolution, vol. 116, Mar. 2018, pp. 1–13.
- ^ a b Copes, Lynn E., and William H. Kimbel. "Cranial Vault Thickness in Primates: Homo Erectus Does Not Have Uniquely Thick Vault Bones." Journal of Human Evolution, vol. 90, Jan. 2016, pp. 120–134.
- ^ Delson E, Harvati K, Reddy D, Marcus LF, Mowbray K, Sawyer GJ, et al. (April 2001). "The Sambungmacan 3 Homo erectus calvaria: a comparative morphometric and morphological analysis". The Anatomical Record. 262 (4): 380–397. doi:10.1002/ar.1048. PMID 11275970. S2CID 25438682.
- ^ Ciochon R, Long VT, Larick R, González L, Grün R, de Vos J, et al. (April 1996). "Dated co-occurrence of Homo erectus and Gigantopithecus from Tham Khuyen Cave, Vietnam". Proceedings of the National Academy of Sciences of the United States of America. 93 (7): 3016–3020. Bibcode:1996PNAS...93.3016C. doi:10.1073/pnas.93.7.3016. PMC 39753. PMID 8610161.
- ^ Schuster AM (September–October 1998). "New Skull from Eritrea". Archaeology. Retrieved 3 October 2015.
- ^ Herries AI, Martin JM, Leece AB, Adams JW, Boschian G, Joannes-Boyau R, et al. (April 2020). "Contemporaneity of Australopithecus, Paranthropus, and early Homo erectus in South Africa". Science. 368 (6486): eaaw7293. doi:10.1126/science.aaw7293. hdl:11568/1040368. PMID 32241925. S2CID 214763272.
Further reading
[edit]- Leakey R, Walker A (November 1985). "Homo Erectus Unearthed". National Geographic. Vol. 168, no. 5. pp. 624–629. ISSN 0027-9358. OCLC 643483454.
- Sigmon BA, Cybulski JS (1981). Homo erectus: Papers in Honor of Davidson Black. University of Toronto Press. JSTOR 10.3138/j.ctvcj2jdw.11.
- Theunissen B, Theunissen LT (2012). Eugène Dubois and the Ape-Man from Java. Springer Netherlands. ISBN 9789400922099.
External links
[edit]- Homo erectus Origins – Exploring the Fossil Record – Bradshaw Foundation
- Archaeology Info Archived 16 May 2011 at the Wayback Machine
- Homo erectus – The Smithsonian Institution's Human Origins Program
- Possible co-existence with Homo Habilis – BBC News
- John Hawks's discussion of the Kocabas fossil
- Peter Brown's Australian and Asian Palaeoanthropology
- The Age of Homo erectus – Interactive Map of the Journey of Homo erectus out of Africa
- Human Timeline (Interactive) – Smithsonian, National Museum of Natural History (August 2016).
| Taxonomy (Hominins) |
| ||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Ancestors |
| ||||||||||||||||||||||||||
| Models |
| ||||||||||||||||||||||||||
| Timelines | |||||||||||||||||||||||||||
| Others | |||||||||||||||||||||||||||
- Homo erectus
- Mammals described in 1892
- Pliocene primates
- Pleistocene primates
- Pleistocene mammals of Africa
- Prehistoric Indonesia
- Prehistoric China
- Prehistoric India
- Prehistoric Kenya
- Prehistoric Tanzania
- Prehistoric Hungary
- Prehistoric Vietnam
- Prehistoric Georgia (country)
- Prehistoric Ethiopia
- Prehistoric Eritrea
- Prehistoric Anatolia
- Prehistoric Spain
- Pleistocene mammals of Asia
- Pleistocene mammals of Europe
- Taxa named by Eugène Dubois
- Tool-using mammals
- Early species of Homo
- Fossil taxa described in 1893