
Hippocampus proper: Difference between revisions
made page to incorporate Regions of hippocampus proper |
ce |
||
| (57 intermediate revisions by 31 users not shown) | |||
| Line 1: | Line 1: | ||
{{Short description|Part of the brain of mammals}} |
|||
{{Use American English|date = March 2019}} |
|||
{{Infobox brain |
{{Infobox brain |
||
| Name = |
| Name = Hippocampus proper |
||
| Latin = |
| Latin = hippocampus proprius, cornu Ammonis |
||
| |
| Image = Hippocampus (brain).jpg |
||
| ⚫ | |||
| GrayPage = 832 |
|||
| Image = HippocampalRegions.jpg |
|||
| ⚫ | |||
| Image2 = CajalHippocampus (modified).png |
| Image2 = CajalHippocampus (modified).png |
||
| Caption2 = Basic circuit of the hippocampus, shown using a modified drawing by Ramon y Cajal. DG: dentate gyrus. Sub: subiculum. EC: entorhinal cortex |
| Caption2 = Basic circuit of the hippocampus, shown using a modified drawing by Ramon y Cajal. DG: dentate gyrus. Sub: subiculum. EC: entorhinal cortex |
||
| BrainInfoType = hier |
|||
| BrainInfoNumber = 165 |
|||
| System = |
| System = |
||
| Artery = |
| Artery = |
||
| Vein = |
| Vein = |
||
| MeshName = |
|||
| MeshNumber = |
|||
}} |
}} |
||
The '''hippocampus proper''' refers to the actual structure of the hippocampus which is made up of four regions or subfields. The subfields use the |
The '''hippocampus proper''' refers to the actual [[structure of the hippocampus]] which is made up of four regions or subfields. The subfields CA1, CA2, CA3, and CA4 use the initials of [[Hippocampus#Name|cornu Ammonis]], an earlier name of the [[hippocampus]]. |
||
==Structure== |
|||
There are four '''hippocampal subfields''', regions in the hippocampus proper which form a [[neural circuit]] called the [[trisynaptic circuit]]. |
|||
===CA1=== |
|||
'''CA1''' is the first region in the hippocampal circuit, from which a major output pathway goes to layer V of the [[entorhinal cortex]]. Another significant output is to the [[subiculum]]. |
|||
===CA2=== |
|||
'''CA2''' is a small region located between CA1 and CA3. It receives some input from layer II of the entorhinal cortex via the [[perforant path]]. Its [[pyramidal cell]]s are more like those in CA3 than those in CA1. It is often ignored due to its small size. |
|||
===CA3=== |
|||
'''CA3''' receives input from the [[mossy fiber (hippocampus)|mossy fiber]]s of the [[granule cell]]s in the [[dentate gyrus]], and also from cells in the entorhinal cortex via the perforant path. The mossy fiber pathway ends in the [[stratum lucidum of hippocampus|stratum lucidum]]. The perforant path passes through the stratum lacunosum and ends in the stratum moleculare. There are also inputs from the [[medial septum]] and from the [[diagonal band of Broca]] which terminate in the stratum radiatum, along with commisural connections from the other side of the hippocampus. |
|||
The pyramidal cells in CA3 have a unique type of dendritic spine called a '''thorny excrescence''' or thorn, only found in CA3 pyramidal cells and hilar mossy cells. The thorn has a thin single spine with a number of heads. Clusters of thorns sit on a dendrite on a broad stem. There are also longer spines called ''long-neck spines''. These unique structures also help to demarcate CA3 from CA2.<ref name="Lauer">{{cite journal |last1=Lauer |first1=M |last2=Senitz |first2=D |title=Dendritic excrescences seem to characterize hippocampal CA3 pyramidal neurons in humans. |journal=Journal of Neural Transmission |date=October 2006 |volume=113 |issue=10 |pages=1469–75 |doi=10.1007/s00702-005-0428-8 |pmid=16465457|s2cid=21440177 }}</ref><ref name="Tsamis">{{cite journal |last1=Tsamis |first1=IK |last2=Mytilinaios |first2=GD |last3=Njau |first3=NS |last4=Fotiou |first4=FD |last5=Glaftsi |first5=S |last6=Costa |first6=V |last7=Baloyannis |first7=JS |title=Properties of CA3 dendritic excrescences in Alzheimer's disease. |journal=Current Alzheimer Research |date=February 2010 |volume=7 |issue=1 |pages=84–90 |doi=10.2174/156720510790274482 |pmid=20205674}}</ref> |
|||
The pyramidal cells in CA3 send some axons back to the dentate gyrus hilus, but they mostly project to regions CA2 and CA1 via the [[Schaffer collateral]]s. There are also a significant number of recurrent connections that terminate in CA3. Both the recurrent connections and the Schaffer collaterals terminate preferentially in the septal area in a dorsal direction from the originating cells. CA3 also sends a small set of output fibers to the lateral septum. |
|||
The region is conventionally divided into three divisions. CA3a is the part of the cell band that is most distant from the dentate (and closest to CA1). CA3b is the middle part of the band nearest to the fimbria and fornix connection. CA3c is nearest to the dentate, inserting into the hilus. CA3 overall, has been considered to be the “pacemaker” of the hippocampus. Much of the synchronous bursting activity associated with interictal epileptiform activity appears to be generated in CA3. Its excitatory collateral connectivity seems to be mostly responsible for this. CA3 uniquely, has pyramidal cell axon collaterals that ramify extensively with local regions and make excitatory contacts with them. CA3 has been implicated in a number of working theories on memory and hippocampal learning processes. Slow oscillatory rhythms (theta-band; 3–8 Hz) are cholinergically driven patterns that depend on coupling of interneurons and pyramidal cell axons via gap junctions, as well as glutaminergic (excitatory) and GABAergic (inhibitory) synapses. Sharp EEG waves seen here are also implicated in memory consolidation.<ref name= "eng">Jerome Engel TAP, ed. Epilepsy: A Comprehensive Textbook in Three Volumes. Philadelphia, PA: Lippincott Williams & Wilkins; 2008</ref> |
|||
A key physiological function of the CA3 is encoding heteroassociative memories using its recurrent circuitry. A seminal hypothesis by John Lisman postulated that during a single theta cycle, a defined set of CA3 principal neurons can activate each other to form a well defined sequence, and the spikes ([[action potentials]]) of these cells tend to coincide with the peaks of the superimposed gamma oscillation.<ref>{{cite journal | vauthors = Jensen O, Lisman JE | title = Hippocampal CA3 region predicts memory sequences: accounting for the phase precession of place cells. | journal = Learning and Memory | volume = 3 | issue = 2 | pages = 279 | date = September 1996 | doi = 10.1101/lm.3.2-3.279 | pmid = 10456097 }}</ref><ref>{{cite journal | vauthors = Lisman JE | title = Relating hippocampal circuitry to function: recall of memory sequences by reciprocal dentate-CA3 interactions. | journal = Neuron | volume = 22 | issue = 2 | pages = 233 | date = February 1999 | doi = 10.1016/s0896-6273(00)81085-5 | pmid = 10069330}}</ref> Approximately a decade later, the existence of well-defined CA3 sequences has experimentally been shown in the laboratory of Loren Frank,<ref>{{cite journal | vauthors = Karlsson MP, Frank LM | title = Awake replay of remote experiences in the hippocampus. | journal = Nature Neuroscience | volume = 12 | issue = 7 | pages = 913 | date = July 2009 | doi = 10.1038/nn.2344 | pmid = 19525943| pmc = 2750914 }}</ref><ref>{{cite journal | vauthors = Carr MF, Karlsson MP, Frank LM | title = Transient slow gamma synchrony underlies hippocampal memory replay. | journal = Neuron | volume = 75 | issue = 4 | pages = 700 | date = August 2012 | doi = 10.1016/j.neuron | pmid = 22920260}}</ref> moreover these results demonstrated that previously encoded sequential experience can be replayed by the CA3 region during episodes called "awake replay". A recent hypothesis postulates that CA3 sequences are built up pair by pair during [[memory encoding]], relying on precisely timed, phase-precessing input from the [[entorhinal cortex]].<ref>{{cite journal | vauthors = Kovács KA | title = Episodic Memories: How do the Hippocampus and the Entorhinal Ring Attractors Cooperate to Create Them? | journal = Frontiers in Systems Neuroscience | volume = 14 | pages = 68 | date = September 2020 | doi = 10.3389/fnsys.2020.559186 | pmid = 33013334 | pmc = 7511719 | doi-access = free }} </ref> This mechanism is based on the synapses of the CA3 recurrent [[axon]] corraterals on the [[dendrites]] of the CA3 population <ref>{{cite journal | vauthors = Bains JS, Longacher JM, Staley KJ | title = Reciprocal interactions between CA3 network activity and strength of recurrent collateral synapses. | journal = Nature Neuroscience | volume = 2 | issue = 8 | pages = 720 | date = August 1999 | doi = 10.1038/11184 | pmid = 10412061}}</ref> that form a complete matrix of connections. |
|||
===CA4=== |
|||
'''CA4''' is a misleading term introduced by Lorente de Nó.<ref name="Lorente de Nó 1934" /> He observed that the pyramidal layer of the CA3 was continuous with polymorphic layer of the dentate gyrus and that the "modified pyramids" (later known as mossy cells<ref name=Amaral1978 />) had Schaffer collaterals similar to CA3 pyramidal cells. Amaral showed that the mossy cells in the CA4 of Lorente de Nó did not have schaffer collaterals and that, in contrast to pyramidal cells, they project to the inner molecular layer of the DG and not to CA1.<ref name=Amaral1978 /> The same author thus concluded that the term CA4 should be abandoned and that the zone should be regarded as the polymorphic layer of the dentate gyrus<ref name=Amaral1978 /> (the area dentata of Blackstad (1956)). The polymorphic layer is often called the hilus or hilar region.<ref name=Amaral2007-1b>{{cite book|vauthors= Anderson P, Morris R, Amaral, Bliss T, O'Keefe J|veditors= Anderson P, Morris R, Amaral, Bliss T, O'Keefe J|title= The hippocampus book|edition= first|year= 2007|publisher= Oxford University Press|location= New York|isbn= 978-0195100273|chapter= The hippocampal formation|chapter-url= https://books.google.com/books?id=zg6oyF1DziQC&pg=PA3|access-date= 2016-12-15|archive-date= 2020-03-15|archive-url= https://web.archive.org/web/20200315125212/https://books.google.com/books?id=zg6oyF1DziQC&pg=PA3|url-status= live}}</ref> The neurons in the polymorphic layer, including mossy cells and GABAergic interneurons, primarily receive inputs from the granule cells in the dentate gyrus in the form of mossy fibers and project to the inner molecular layer of the dentate gyrus via the associational/commissural projection.<ref name="Lorente de Nó 1934">{{cite journal |
|||
| last = Lorente de Nó |
|||
| first = R |
|||
| title = Studies on the structure of the cerebral cortex. Continuation of the study of the ammonic system |
|||
| journal = J. Psychol. Neurol. |
|||
| volume = 46 |
|||
| pages = 113–177 |
|||
| date = 1934 |
|||
| ref = Lorente1934 |
|||
}}</ref><ref name=Amaral1978>{{cite journal |
|||
| last = Amaral |
|||
| first = DG |
|||
| title = A Golgi study of cell types in the hilar region of the hippocampus in the rat |
|||
| journal = J. Comp. Neurol. |
|||
| volume = 182 |
|||
| pages = 851–914 |
|||
| date = 1978 |
|||
| pmid = 730852 |
|||
| doi = 10.1002/cne.901820508 |
|||
| issue = 5 |
|||
| s2cid = 44257239 |
|||
}}</ref> |
|||
They also receive a small number of connections from pyramidal cells in CA3. They, in turn, project back into the dentate gyrus at distant septotemporal levels. |
|||
==Additional images== |
|||
<gallery> |
|||
File:Coronary brain section through hippocampus.jpg|High resolution micrograph of human brain, [[coronal section]] through the hippocampus. |
|||
File:HippocampalRegions.jpg|Diagram of hippocampal regions in a rat brain. |
|||
</gallery> |
|||
==References== |
|||
{{Reflist}} |
|||
{{Rostral basal ganglia and associated structures}} |
|||
{{Papez circuit}} |
|||
{{Authority control}} |
|||
[[Category:Hippocampus (brain)]] |
|||
Latest revision as of 15:24, 13 November 2024
| Hippocampus proper | |
|---|---|
.jpg) Hippocampus location and regions in a human, coronal plane | |
.png) Basic circuit of the hippocampus, shown using a modified drawing by Ramon y Cajal. DG: dentate gyrus. Sub: subiculum. EC: entorhinal cortex | |
| Details | |
| Identifiers | |
| Latin | hippocampus proprius, cornu Ammonis |
| NeuroNames | 182 |
| TA98 | A14.1.09.327 |
| TA2 | 5520 |
| FMA | 62493 |
| Anatomical terms of neuroanatomy | |
The hippocampus proper refers to the actual structure of the hippocampus which is made up of four regions or subfields. The subfields CA1, CA2, CA3, and CA4 use the initials of cornu Ammonis, an earlier name of the hippocampus.
Structure
[edit]There are four hippocampal subfields, regions in the hippocampus proper which form a neural circuit called the trisynaptic circuit.
CA1
[edit]CA1 is the first region in the hippocampal circuit, from which a major output pathway goes to layer V of the entorhinal cortex. Another significant output is to the subiculum.
CA2
[edit]CA2 is a small region located between CA1 and CA3. It receives some input from layer II of the entorhinal cortex via the perforant path. Its pyramidal cells are more like those in CA3 than those in CA1. It is often ignored due to its small size.
CA3
[edit]CA3 receives input from the mossy fibers of the granule cells in the dentate gyrus, and also from cells in the entorhinal cortex via the perforant path. The mossy fiber pathway ends in the stratum lucidum. The perforant path passes through the stratum lacunosum and ends in the stratum moleculare. There are also inputs from the medial septum and from the diagonal band of Broca which terminate in the stratum radiatum, along with commisural connections from the other side of the hippocampus.
The pyramidal cells in CA3 have a unique type of dendritic spine called a thorny excrescence or thorn, only found in CA3 pyramidal cells and hilar mossy cells. The thorn has a thin single spine with a number of heads. Clusters of thorns sit on a dendrite on a broad stem. There are also longer spines called long-neck spines. These unique structures also help to demarcate CA3 from CA2.[1][2]
The pyramidal cells in CA3 send some axons back to the dentate gyrus hilus, but they mostly project to regions CA2 and CA1 via the Schaffer collaterals. There are also a significant number of recurrent connections that terminate in CA3. Both the recurrent connections and the Schaffer collaterals terminate preferentially in the septal area in a dorsal direction from the originating cells. CA3 also sends a small set of output fibers to the lateral septum.
The region is conventionally divided into three divisions. CA3a is the part of the cell band that is most distant from the dentate (and closest to CA1). CA3b is the middle part of the band nearest to the fimbria and fornix connection. CA3c is nearest to the dentate, inserting into the hilus. CA3 overall, has been considered to be the “pacemaker” of the hippocampus. Much of the synchronous bursting activity associated with interictal epileptiform activity appears to be generated in CA3. Its excitatory collateral connectivity seems to be mostly responsible for this. CA3 uniquely, has pyramidal cell axon collaterals that ramify extensively with local regions and make excitatory contacts with them. CA3 has been implicated in a number of working theories on memory and hippocampal learning processes. Slow oscillatory rhythms (theta-band; 3–8 Hz) are cholinergically driven patterns that depend on coupling of interneurons and pyramidal cell axons via gap junctions, as well as glutaminergic (excitatory) and GABAergic (inhibitory) synapses. Sharp EEG waves seen here are also implicated in memory consolidation.[3]
A key physiological function of the CA3 is encoding heteroassociative memories using its recurrent circuitry. A seminal hypothesis by John Lisman postulated that during a single theta cycle, a defined set of CA3 principal neurons can activate each other to form a well defined sequence, and the spikes (action potentials) of these cells tend to coincide with the peaks of the superimposed gamma oscillation.[4][5] Approximately a decade later, the existence of well-defined CA3 sequences has experimentally been shown in the laboratory of Loren Frank,[6][7] moreover these results demonstrated that previously encoded sequential experience can be replayed by the CA3 region during episodes called "awake replay". A recent hypothesis postulates that CA3 sequences are built up pair by pair during memory encoding, relying on precisely timed, phase-precessing input from the entorhinal cortex.[8] This mechanism is based on the synapses of the CA3 recurrent axon corraterals on the dendrites of the CA3 population [9] that form a complete matrix of connections.
CA4
[edit]CA4 is a misleading term introduced by Lorente de Nó.[10] He observed that the pyramidal layer of the CA3 was continuous with polymorphic layer of the dentate gyrus and that the "modified pyramids" (later known as mossy cells[11]) had Schaffer collaterals similar to CA3 pyramidal cells. Amaral showed that the mossy cells in the CA4 of Lorente de Nó did not have schaffer collaterals and that, in contrast to pyramidal cells, they project to the inner molecular layer of the DG and not to CA1.[11] The same author thus concluded that the term CA4 should be abandoned and that the zone should be regarded as the polymorphic layer of the dentate gyrus[11] (the area dentata of Blackstad (1956)). The polymorphic layer is often called the hilus or hilar region.[12] The neurons in the polymorphic layer, including mossy cells and GABAergic interneurons, primarily receive inputs from the granule cells in the dentate gyrus in the form of mossy fibers and project to the inner molecular layer of the dentate gyrus via the associational/commissural projection.[10][11] They also receive a small number of connections from pyramidal cells in CA3. They, in turn, project back into the dentate gyrus at distant septotemporal levels.
Additional images
[edit]-
 High resolution micrograph of human brain, coronal section through the hippocampus.
High resolution micrograph of human brain, coronal section through the hippocampus. -
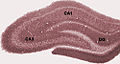 Diagram of hippocampal regions in a rat brain.
Diagram of hippocampal regions in a rat brain.


References
[edit]- ^ Lauer, M; Senitz, D (October 2006). "Dendritic excrescences seem to characterize hippocampal CA3 pyramidal neurons in humans". Journal of Neural Transmission. 113 (10): 1469–75. doi:10.1007/s00702-005-0428-8. PMID 16465457. S2CID 21440177.
- ^ Tsamis, IK; Mytilinaios, GD; Njau, NS; Fotiou, FD; Glaftsi, S; Costa, V; Baloyannis, JS (February 2010). "Properties of CA3 dendritic excrescences in Alzheimer's disease". Current Alzheimer Research. 7 (1): 84–90. doi:10.2174/156720510790274482. PMID 20205674.
- ^ Jerome Engel TAP, ed. Epilepsy: A Comprehensive Textbook in Three Volumes. Philadelphia, PA: Lippincott Williams & Wilkins; 2008
- ^ Jensen O, Lisman JE (September 1996). "Hippocampal CA3 region predicts memory sequences: accounting for the phase precession of place cells". Learning and Memory. 3 (2): 279. doi:10.1101/lm.3.2-3.279. PMID 10456097.
- ^ Lisman JE (February 1999). "Relating hippocampal circuitry to function: recall of memory sequences by reciprocal dentate-CA3 interactions". Neuron. 22 (2): 233. doi:10.1016/s0896-6273(00)81085-5. PMID 10069330.
- ^ Karlsson MP, Frank LM (July 2009). "Awake replay of remote experiences in the hippocampus". Nature Neuroscience. 12 (7): 913. doi:10.1038/nn.2344. PMC 2750914. PMID 19525943.
- ^ Carr MF, Karlsson MP, Frank LM (August 2012). "Transient slow gamma synchrony underlies hippocampal memory replay". Neuron. 75 (4): 700. doi:10.1016/j.neuron. PMID 22920260.
- ^ Kovács KA (September 2020). "Episodic Memories: How do the Hippocampus and the Entorhinal Ring Attractors Cooperate to Create Them?". Frontiers in Systems Neuroscience. 14: 68. doi:10.3389/fnsys.2020.559186. PMC 7511719. PMID 33013334.
- ^ Bains JS, Longacher JM, Staley KJ (August 1999). "Reciprocal interactions between CA3 network activity and strength of recurrent collateral synapses". Nature Neuroscience. 2 (8): 720. doi:10.1038/11184. PMID 10412061.
- ^ a b Lorente de Nó, R (1934). "Studies on the structure of the cerebral cortex. Continuation of the study of the ammonic system". J. Psychol. Neurol. 46: 113–177.
- ^ a b c d Amaral, DG (1978). "A Golgi study of cell types in the hilar region of the hippocampus in the rat". J. Comp. Neurol. 182 (5): 851–914. doi:10.1002/cne.901820508. PMID 730852. S2CID 44257239.
- ^ Anderson P, Morris R, Amaral, Bliss T, O'Keefe J (2007). "The hippocampal formation". In Anderson P, Morris R, Amaral, Bliss T, O'Keefe J (eds.). The hippocampus book (first ed.). New York: Oxford University Press. ISBN 978-0195100273. Archived from the original on 2020-03-15. Retrieved 2016-12-15.