Фотосистема I
Фотосисте́ма I (первая фотосистема, фотосистема один, ФСI), или пластоциани́н-ферредокси́н-оксидоредукта́за — второй функциональный комплекс электрон-транспортной цепи (ЭТЦ) хлоропластов. Он принимает электрон от пластоцианина и, поглощая световую энергию, формирует сильный восстановитель П700, способный через цепь переносчиков электронов осуществить восстановление НАДФ+. Таким образом, при участии ФСI синтезируется источник электронов (НАДФН) для последующих реакций восстановления углерода в хлоропластах в цикле Кальвина. Кроме того, ФСI может осуществлять циклический транспорт электронов, сопряжённый с синтезом АТФ, обеспечивая дополнительный синтез АТФ в хлоропластах[1].
История открытия
Нециклический транспорт электронов начинается с того, что марганцевый кластер фотосистемы II окисляет воду, восстанавливая пул пластохинонов. Далее цитохром b6f-комплекс окисляет пластохиноны, а электрон через пластоцианин передаётся на фотосистему I, где используется для синтеза НАДФН. Нарушение формальной логики в названиях фотосистем связано с тем, что фотосистема I была открыта раньше, чем фотосистема II.
Первые данные указывающие на существование ФСI появились в 1950-х годах, но в то время никто ещё не мог оценить значимость этих открытий[2]. Представление о существовании в хлоропластах двух фотосистем возникло в уже в 1940-е годы на основание опытов лаборатории Р. Эмерсона, обнаружившего эффект падения квантового выхода фотосинтеза при освещении хлоропластов монохроматическим красным светом (λ>680 нм), возбуждающим только ФСI, и эффект усиления квантового выхода при добавлении подсветки с длиной волны около 650 нм, которая возбуждала ФСII (так называемый эффект Эмерсона). Также следует упомянуть открытый Комоннером в 1956 году светоиндуцируемый ЭПР сигнал, который был назван сигналом I. По чистой случайности сигнал I и сигнал II, исходили от ФСI и ФСII соответственно[2]. Лишь в 1960 году Луис Дюйзенс предложил концепцию фотосистемы I и фотосистемы II, и в том же году Фэй Бендалл и Роберт Хилл организовали результаты предыдущих открытий в стройную теорию последовательных реакций фотосинтеза[2]. Гипотеза Хилла и Бендалля была позднее подтверждена в экспериментах Дюйзенса и Витта в 1961 году[2].
После этого начались систематические попытки физического выделения фотосистемы I, определения её трёхмерной структуры и тонкого строения. В 1966 году начался бум исследований в этой области: Андерсон и Бордман подвергали мембраны хлоропластов воздействию ультразвука с последующей обработкой дигитонином, Вернон использовал тритон X-100, а Огава — додецилсульфат. Однако, первые полученные экстракты содержали примеси светосособирающих комплексов, а также цитохромов f и b6. Потребовалось много времени что бы разобраться, что полученные экстракты были смесью[2].
В 1968 году Рид и Клайтон смогли выделить реакционный центр фотосистемы I из пурпурных бактерий, что значительно подстегнуло исследования оксигенного фотосинтеза. Однако, открытым оставался вопрос: что из выделенного являлось истинным реакционным центром, что антенными комплексами, а что дополнительными субъединицами. Долгое время эффективное выделение реакционного центра фотосистемы I оставалось неразрешённой проблемой. В конце-концов оказалось, что легче всего это сделать у цианобактерий, поскольку у них отсутствовали интегрированные в мембрану внешние антенны. После многочисленных попыток с разными вида, выяснилось, что самыми многообещающими в этом отношение видами являются представителями Synechocystis и Synechococcus, поскольку фотосистема I, выделенная из Thermosynechococcus elongatus давала очень стабильный реакционный центр, пригодный для кристаллизации и исследования методом рентгеноструктурного анализ[2].
Отличия от фотосистемы II
Основная функция фотосистемы II — генерация сильного окислителя, который инициирует окисление воды и передачу её электронов на мембранный переносчик. Основная функция фотосистемы I — насытить эти низкоуровневые электроны энергией, что бы с их помощью осуществить восстановление НАДФ+. Поскольку энергия суммарного процесса слишком велика, что бы осуществить его в рамках одного реакционного центра, в ходе эволюции появились две фотосистемы, которые раздельно осуществляют разные части этой реакции. Их специфические функции и определяют особенности их строения. Так, фотосистема I — симметрична, то есть в ней работают две ветви электронного транспорта, что делает его значительно более быстрым, в то время как фотосистема II — асимметрична и обладает только одной рабочей ветвью, что замедляет транспорт электронов, но делает его более управляемым. Обе фотосистемы значительно отличаются по строению антенн, дополнительных субъединиц, способов регуляции и своему положению в мембране[3]. Так, фотосистема I обладает интегральной антенной, хлорофиллы которой расположены непосредственно на главных белках комплекса — А и B, в то время как у фотосистемы II они вынесены на внешние белки CP47 и CP43. По количеству дополнительных малых регуляторных субъединиц ФС II значительно превосходит ФС I, что связано с необходимостью тонкой регуляции процесса окисления воды, который потенциально крайне опасен для клетки. Этим же объясняется не однородное распределение фотосистем в мембране тилакоида: в то время как ФС I располагается преимущественно в области маргинальных, торцевых и стромальных мембран, ФС II практически полностью находится в области спаренных мембран, что обеспечивает клетке дополнительную защиту от продуцируемых ею активных форм кислорода[4].
Главное отличие фотосистемы II от фотосистемы I — это наличие большого обращённого в люмен домена, который содержит марганцевый кластер и окружающие его защитные белки. Именно здесь происходит процесс фотохимического окисления воды, сопровождаемый выделением кислорода и протонов[3].
Структурная организация фотосистемы I
| Фотосистема I | |
|---|---|
 Растительная Фотосистема I | |
| Идентификаторы | |
| Шифр КФ | 1.97.1.12 |
| Базы ферментов | |
| IntEnz | IntEnz view |
| BRENDA | BRENDA entry |
| ExPASy | NiceZyme view |
| MetaCyc | metabolic pathway |
| KEGG | KEGG entry |
| PRIAM | profile |
| PDB structures | RCSB PDB PDBe PDBj PDBsum |
| Поиск | |
| PMC | статьи |
| PubMed | статьи |
| NCBI | NCBI proteins |
| PsaA_PsaB | |
|---|---|
 Структура Фотосистемы I из цианобактерии. Показана интегральная часть(ядро) и белки внешней антенны. | |
| Идентификаторы | |
| Символ | PsaA_PsaB |
| Pfam | PF00223 |
| InterPro | IPR001280 |
| PROSITE | PDOC00347 |
| SCOP | 1jb0 |
| SUPERFAMILY | 1jb0 |
| TCDB | 5.B.4 |
| OPM superfamily | 2 |
| OPM protein | 1jb0 |
| Доступные структуры белков | |
| Pfam | структуры |
| PDB | RCSB PDB; PDBe; PDBj |
| PDBsum | 3D-модель |
Фотосистема I состоит из следующих белковых субъединиц и кофакторов[5][6][1]:
| Субъединицы | Описание |
|---|---|
| A | 83 кДа, 751 аминокислотных остатков |
| B | 82,5 кДа, 735 аминокислотных остатков |
| C | 8,9 кДа, перенос электрона с П700 на ферредоксин |
| D | 19 кДа, обеспечивает связь с ферредоксином |
| E | 7,5 кДа, обеспечивает связь с ферредоксином |
| F | 19 кДа, взаимодействует с пластоцианином |
| G | 8 кДа, только у растений |
| H | 10 кДа, у растений предотвращает формирования тримеров ФСI, обеспечивает взаимодействие со светособирающим комплексом II |
| I | 5 кДа, у растений взаимодействует с PsaH, связывается со светособирающим комплексом II; у цианобактерий играет важную роль в формировании тримеров ФСI |
| J | 5 кДа, несёт три молекулы хлорофилла и выполняет структурную функцию |
| K | 8.5 кДа, несёт две молекулы хлорофилла и выполняет структурную функцию |
| L | 16 кДа, у цианобактерий участвуют в формировании тримера ФСI; у растений она связывает со светособирающим комплексом II |
| M | 3,5 кДа, только у цианобактерий; расположена в области контакта ФСI в тримере |
| N | 9 кДа, есть у растений и водорослей |
| O | только у растений; функция неизвестна |
| X | 4 кДа, только у цианобактерий |
| Пигменты | |
| Хлорофилл a | 95 молекул в антенной системе |
| Хлорофилл а | 2 молекулы дополнительного хлорофилла а |
| Хлорофилл a0 | Хлорофилл а695 — первичный акцептор электронов |
| Хлорофиллы а и a' | специальная пара П700 |
| β-Каротин | 22 молекулы |
| Коферменты/Кофакторы | |
| Fa | Fe4S4 железосерный кластер (ЭТЦ) |
| Fb | Fe4S4 железосерный кластер (ЭТЦ) |
| Fx | Fe4S4 железосерный кластер (ЭТЦ) |
| Ферредоксин | Переносчик электронов |
| Пластоцианин | Растворимый белок, содержащий атом меди |
| QK-A | Филлохинон — акцептор электронов в ЭТЦ (субъединица А) |
| QK-B | Филлохинон — акцептор электронов в ЭТЦ (субъединица В) |
| Ca2+ | ион кальция |
| Mg2+ | ион магния |
Основная функция ФСI — передача энергии света на электрон, перенос электрона от пластоцианина к ферредоксину[7]. ФСI содержит свыше 110 кофакторов, значительно больше чем фотосистема II[8]. Каждый из этих компонентов имеет широкий спектр функций. Основные компоненты электрон-транспортной цепи ФСI — главный донор возбуждённых электронов П700 (хлорофильный димер) и пять переносчиков: A0 (хлорофилл а), A1 (филлохинон) и три Fe4S4 железосерных кластера: Fx, Fa, и Fb[9].
Структурно ФСI представляет собой гетеродимер двух интегральных белковых комплексов — А и В (у всех растений кодируются хлоропластными генами PsaA и PsaB). Белки А и В присоединяют димер Р700, по одной молекуле мономера хлорофилла а (Хл695) — первичного акцептора электронов А0, по одному дополнительному хлорофиллу а и по одной молекуле филлохинона (А1). Два набора дополнительных хлорофиллов а, первичных акцепторов электрона и филлохинонов формируют две почти симметричные ветви транспорта электронов от Р700 к Fx. В отличие от реакционных центров зелёных и пурпурных бактерий и ФСII, где из двух ветвей функционирует лишь одна, в ФСI активны обе ветви электронного транспорта, хотя они и не идентичны[1].Белок А гомологичен белкам D1+СP43 (молекулярная масса белка А соответствует сумме молекулярных масс белков D1 и СP43) из фотосистемы II, а белок В гомологичен белкам D2+CP47 соответственно[10].
Обе субъединицы содержат 11 трансмембранных сегментов[англ.]. Железосодержащий кластер Fx связан четырьмя цистеинами, два из которых находятся на субъединице А, и ещё два на субъединице В. В обоих белках цистеины находятся на проксимальном конце, в петле между девятым и десятым трансмембранными сегментами. По всей вероятности, ниже цистеинов находится так называемый мотив лейциновой молнии, который вносит существенный вклад в димеризацию белков А и В[11]. Конечные акцепторы электронов FA и FB находятся на субъединице С[12][13].
Следует особо подчеркнуть, что перенос электрона осуществляется в соответствии с термодинамическим потенциалом. Увеличение окислительно-восстановительных потенциалов в цепи акцепторов обеспечивает быстрое снижение энергии, что предотвращает возврат электрона к пигменту и бесполезную трату энергии электронного возбуждения. Благодаря этому энергия возбуждения эффективно используется для разделения зарядов[14].
Пластоцианин

Пластоцианин — маленький, подвижный белок с молекулярной массой около 10,5 кДа. К его центральному атому Сu присоединяются остатки цистеина и метионина, а сбоку его стабилизируют два остатка гистидина. При обратимой смене валентности Cu2+ ↔ Cu+1 пластоцианин либо поглощает один электрон, либо отдаёт его. Пластоцианин является аналогом цитохрома c, который выполняет схожую функцию в дыхательной цепи митохондрий[6].
Он принимает электрон от цитохром b6f-комплекса, окисляя цитохром f и перенося его непосредственно на реакционный центр П700 фотосистемы I. На внешней стороне белка имеется группа аминокислот, несущих отрицательный заряд[16]. Предположительно, они связываются с положительно заряженным люминальным доменом субъединицы F, однако механизм связывания недостаточно изучен и остаётся неясным[17].
У некоторых водорослей и цианобактерий при недостатке меди в среде пластоцианин не образуется, вместо него синтезируется и выполняет его функции цитохром c-553[18].
- Пластоцианин (PC) отдаёт один электрон окисленному П700+ и восстанавливает его до исходного состояния:

Специальная пара П700
П700 (в английской литературе P700) представляет собой димер хлорофилла а и хлорофилла а‘ у которого кетоэфирная группа в V кольце находится в цис-положении относительно плоскости молекулы, с максимумом поглощения 700 нм[19]. Наличие цис-кетоэфирной группы делает возможным образование димера из двух хлорофиллов посредством формирования водородных связей. П700 получает энергию от антенных комплексов и использует её, чтобы поднять электроны на более высокий уровень. Далее электрон в ходе окислительно-восстановительной реакции переходит на цепь переносчиков. В окисленном состоянии окислительно-восстановительный редокс-потенциал П700 составляет +0,52 В, а в фотовозбуждённом состоянии он становится −1,2 В, то есть формируется мощный восстановитель, обеспечивающий восстановление НАДФ+[20][21].
- В соответствии со следующим уравнением П700 поглощает квант света и переходит в фотовозбуждённое состояние, в результате чего один из его электронов переходит с основного подуровня S0 на первый синглетный подуровень S1:

Хлорофилл А0
A0 — первый акцептор электронов в фотосистеме I. Именно здесь происходит первичное фотохимическое разделение зарядов между фотовозбуждённым П700* и A0. Его максимум поглощения составляет 695 нм (Хл а695), что объясняется его взаимодействием с окружающими аминокислотными остатками[19]. Его редокс-потенциал в восстановленном состоянии −1,1 В[1].
- Фотовозбуждённый П700* отдаёт один электрон хлорофиллу A0, в результате чего происходит разделение зарядов, и образуется первичная радикальная пара:

Филлохинон A1
Следующий акцептор — это Филлохинон A1, также известный как витамин K1. Он, как и хлорофилл, обладает фитольным хвостом[22], и приблизительно соответствует пластохинону QA фотосистемы II. Поглощая электрон, он образует семихинон-радикал, который восстанавливает Fx, передаёт его на Fb и далее на Fa[22][23].

Железосерные кластеры
Железосерные кластеры ФСI имеют форму куба с четырьмя атомами железа и четырьмя атомами серы, составляющими его восемь вершин. Все три кластера связаны с белками ФСI через остатки цистеина[24]. Fx (Eо‘ = −0,70 В) окисляет восстановленный А1. Дальнейший транспорт осуществляют железосерные кластеры Fa, и Fb, характеризующиеся низкими окислительно-восстановительными потенциалами (-0,59 и −0,55 В соответственно). Множество экспериментов выявило несоответствие между разными теориями, описывающими расположение и работу железосерных кластеров[24]. Однако большинство результатов позволяет сделать некоторые общие выводы. Во-первых, Fx, Fa, и Fb образуют треугольник, и Fa располагается ближе к Fx чем Fb[24]. Во-вторых, транспорт электронов начинается с Fx через Fa к Fb, или же через Fa к Fb. До сих пор ведутся споры о том, какой из двух кластеров осуществляет перенос электрона на ферредоксин[24].
Ферредоксин
Ферредоксин — это водорастворимый белок с молекулярной массой 11 кДа и содержащий Fe2S2 центр[25]. Примечательно, что он является одноэлектронной окислительно-восстановительной системой, то есть переносит только один электрон, полученный им от железосерных кластеров. Он восстанавливается ФСI на стромальной стороне мембраны и в восстановленном состоянии является сильным восстановителем (Eo‘ = —0,6 В), благодаря чему может быть переносчиком электронов для различных реакций, протекающих в хлоропласте. Так, ферредоксин поставляет электроны для восстановления нитритов, (нитритредуктаза) и ассимиляции серы (сульфитредуктаза) в хлоропласте. Также он поставляет электроны для фиксации атмосферного азота (нитрогеназа) у бактерий. Он восстанавливает тиоредоксин — низкомолекулярный серосодержащий белок, участвующий в редокс-регуляции хлоропластов, активируя ключевые ферменты цикла Кальвина. При нециклическом транспорте электронов ферредоксин взаимодействует с ферредоксин-НАДФ+-редуктазой, которая восстанавливает НАДФ+ до НАДФН (Eo‘ = −0,32 В) в строме хлоропласта[25].
Светособирающий комплекс

Светособирающие комплексы состоят из молекул хлорофилла а и b и каротиноидов, соединённых с белками[20]. Эти пигменты, возбуждаясь, переносят энергию фотонов в реакционный центр фотосистемы по Фёрстеровскому механизму. В отличие от реакционного центра ФСI, светособирающие комплексы могут поглощать практически во всей области видимого спектра[26]. Антенные комплексы подразделяют на внутренние, или интегральные, антенны, непосредственно присоединённые к комплексу фотосистемы, и периферические подвижные светособирающие комплексы (ССКI). Так, белки А и В присоединяют пигменты внутренней антенны ФСI: около 95 молекул хлорофилла а и 22 молекулы β-каротина, 5 из которых находятся в цис-конформации. В координации по крайней мере десяти хлорофиллов внутренней антенны принимают участие малые субъединицы J, K, L, M и X. Пигменты внутренней антенны располагаются в виде цилиндра, окружающего все компоненты электрон-транспортной цепи фотосистемы I. Это отличает ФСI от ФСII, где пигменты внутренней антенны расположены на отдельных белках CP43 и CP77[1]. Внешний светособирающий комплекс ССКI (LHCI) содержит 80—120 молекул хлорофиллов а и b, каротиноиды и состоит из четырёх субъединиц: Lhca1, Lhca2, Lhca3 и Lhca4 — с молекулярными массами 17—24 кДа. Относительно недавно были открыты две дополнительные субъединицы, Lhca5 и Lhca6, однако их концентрация в мембране тилакоида крайне мала, а кодирующие их гены практически не экспрессируются[27][28].
Циклический транспорт электронов

При слишком сильном освещении и/или закрытых устьицах (голодание по СО2) происходит перевосстановление пула пластохинонов и, как следствие, перевосстановление пула НАДФ+. При недостатке СО2 НАДФН не может расходоваться в цикле Кальвина, а значит, не хватает субстрата для ферредоксин-НАДФ+-редуктазы[англ.]. В конечном итоге это приводит к тому, что ФСI становится некуда сбрасывать возбуждённые электроны, а это в свою очередь может привести к повреждению фотосинтетического аппарата, окислению мембран и образованию активных форм кислорода[6]. В этих условиях, чтобы предотвратить окислительный стресс и защититься от фотоповреждения, растения переходят к циклическому транспорту электронов. Полагают, что катализатором циклического транспорта является восстановленный ферредоксин[29][30].
Циклическое фотофосфорилирование
Вначале электрон неким образом перемещается от восстановленного ферредоксина на пул пластохинонов. Точный механизм этого процесса не известен. Полагают, что эту реакцию осуществляет особый фермент — ферредоксин-пластохинон-оксидоредуктаза. Затем от пластохинона через цитохром b6f-комплекс и пластоцианин электрон вновь попадает на ФСI. При этом происходит закачка протона в полость тилакоида и идёт синтез АТФ. В качестве наиболее вероятного кандидата на роль ферредоксин-пластохинон-оксидоредуктазы в последнее время рассматривают ферредоксин-НАДФ+-редуктазу, которая может образовывать комплекс с цитохром-b6f-комплексом. Предположительно она может переносить электроны с ферредоксины напрямую на убихинон, связанный цитохром-b6f-комплексом через специальный гем cn[31][32]. Большое количество данных также говорит в пользу образования суперкомплекса из цитохром-b6f-комплекса, ФСI, ферредоксин-НАДФ+-редуктазы и трансмембранного белка PGRL1. Образование и распад такого комплекса, как полагают, переключает режим потока электрона с нециклического на циклический и обратно[33][34].
Ещё один фермент, возможно принимающей участие а этом процесс — это НАДН-дегидрогеназный комплекс хлоропластов, аналогичный НАДH-дегидрогеназному комплексу митохондрий и гомологичный бактериальному комплексу I[35][36]. Он окисляет ферредоксин и сбрасывает электроны на пластохинон, предотвращая окислительный стресс. НАДН-дегидрогеназный комплекс хлоропластов образует суперкомплекс с двумя ФСI при помощи белков Lhca5 и Lhca6[28]. Протонный градиент, созданный в результате циклического фтофосфорилирования на мембране тилакоида, используется белками-переносчиками для встраивания в мембрану приходящих из стромы белков[37][38].
Псевдоциклический транспорт
При очень активном восстановлении пула ферредоксинов происходит сброс их электронов на О2 с образованием H2O (так называемая реакция Мелера). Он сходен с циклическим транспортом тем, что не синтезируется НАДФН, а только АТФ. Однако в условиях реакции Мелера соотношение АТФ/АДФ очень велико, так что имеющегося количества АДФ не достаточно для синтеза АТФ, и, как следствие, на мембране тилакоида создаётся очень высокий протонный градиент. В результате реакции происходит образование супероксид-анион-радикала O2-·, который превращается в O2 и H2O2 под воздействием фермента супероксиддисмутазы, а перекись превращается в воду ферментом аскорбатпероксидазой[6].

Ещё один фермент, участвующий в псевдоциклическом транспорте, это терминальная оксидаза хлоропластов, гомологичная альтернативной оксидазе растительных митохондрий. Она окисляет пул пластохинонов с участием кислорода, образуя воду и рассеивая энергию в форме тепла[39].
Локализация в мембране тилакоида
Фотосистема I находится в стромальных тилакоидах (32 %), а также в маргинальных (36 %) и торцевых (32 %) областях гран. Такое расположение обусловлено плотностью её поверхностного заряда и силами электростатического отталкивания с другими комплексами[40].
У цианобактерий и прохлорофитов фотосистема I способна образовывать тримеры. Это способствует увеличению спектра поглощения на больших глубинах, а также более эффективному перераспределению энергии возбуждения и защите от фотоповреждений[41]. Следует отметить, что у эукариот фотосистема I утратила эту способность благодаря наличию субъединицы H, а также мутации в субъединице L. Вместо тримерезации у эукариот она при помощи субъединиц L и G взаимодействует с большими мембранными светособирающими комплексами, которых нет у прокариот[42].
Белок Ycf4
Трансмембранный белок Ycf4, обнаруженный в мембране тилакоида, жизненно необходим для функционирования фотосистемы I. Он участвует в сборке компонентов комплекса, без него фотосинтез становится неэффективным[43].
Зелёные серобактерии и эволюция ФСI
Молекулярно-биологические данные говорят в пользу того, что ФСI, вероятно, эволюционировала из фотосистемы зелёных серобактерий. Реакционные центры зелёных серобактерий, цианобактерий, водорослей и высших растений различаются, однако домены, выполняющие аналогичные функции, имеют схожее строение[44]. Так, во всех трёх системах окислительно-восстановительный потенциал достаточен для восстановления ферредоксина[44]. Все три электронотранспортные цепи содержат железо-серные белки[44]. И наконец, все три фотосистемы представляют собой димер двух гидрофобных белков, на которых закреплены редокс-центры и пигменты интегральной антенны[44]. В свою очередь, фотосистема зелёных серобактерий содержит те же самые кофакторы, что и электронотранспортная цепь фотосистемы I[44].
Галерея
-
 Положение хлорофиллов и кофакторов в фотосистеме I.
Положение хлорофиллов и кофакторов в фотосистеме I. -
 Тример фотосистемы I
Тример фотосистемы I -
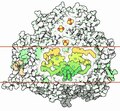 ЭТЦ фотосистемы I
ЭТЦ фотосистемы I -
 Фотосистемы I и реакционный центр бактерии.
Фотосистемы I и реакционный центр бактерии. -
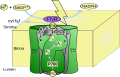 Модель фотосистемы I.
Модель фотосистемы I.


{kind=link}
{kind=link}
{kind=link}
См. также
Примечания
- ↑ 1 2 3 4 5 Ермаков, 2005, с. 173-175.
- ↑ 1 2 3 4 5 6 Fromme P, Mathis P (2004). "Unraveling the photosystem I reaction center: a history, or the sum of many efforts" (PDF). Photosyn. Res. 80 (1–3): 109—24. doi:10.1023/B:PRES.0000030657.88242.e1. PMID 16328814.
- ↑ 1 2 Ермаков, 2005, с. 121.
- ↑ Ravi Danielsson, Marjaana Suorsa, Virpi Paakkarinen, Per-Åke Albertsson, Stenbjörn Styring, Eva-Mari Aro and Fikret Mamedov (2006). "Dimeric and Monomeric Organization of Photosystem II". The Journal of Biological Chemistry (281): 14241–14249. doi:10.1074/jbc.M600634200.
{{cite journal}}: Неизвестный параметр|month=игнорируется (справка)Википедия:Обслуживание CS1 (множественные имена: authors list) (ссылка) Википедия:Обслуживание CS1 (не помеченный открытым DOI) (ссылка) - ↑ Saenger W, Jordan P, Krauss N (April 2002). "The assembly of protein subunits and cofactors in photosystem I". Curr. Opin. Struct. Biol. 12 (2): 244—54. doi:10.1016/S0959-440X(02)00317-2. PMID 11959504.
{{cite journal}}: Википедия:Обслуживание CS1 (множественные имена: authors list) (ссылка) - ↑ 1 2 3 4 Страсбургер, 2008, с. 117.
- ↑ Golbeck JH (1987). "Structure, function and organization of the Photosystem I reaction center complex". Biochim. Biophys. Acta. 895 (3): 167—204. doi:10.1016/s0304-4173(87)80002-2. PMID 3333014.
- ↑ HongQi Yu', Ingo Gortjohann, Yana Bukman, Craig Yolley', Devendra K. Chauhan, Alexander Melkozerov and Petra Fromme. "Structure and funcnions of photosystems I and II" (PDF).
{{cite journal}}: Cite journal требует|journal=(справка)Википедия:Обслуживание CS1 (множественные имена: authors list) (ссылка) - ↑ Jagannathan, Bharat; Golbeck, John (2009). "Photosynthesis:Microbial". Encyclopedia of Microbiology, 3rd Ed: 325—341. doi:10.1016/B978-012373944-5.00352-7.
{{cite journal}}: Википедия:Обслуживание CS1 (множественные имена: authors list) (ссылка) - ↑ Хелдт, 2011, с. 99.
- ↑ Webber AN, Malkin R (May 1990). "Photosystem I reaction-centre proteins contain leucine zipper motifs. A proposed role in dimer formation". FEBS Lett. 264 (1): 1—4. doi:10.1016/0014-5793(90)80749-9. PMID 2186925.
- ↑ Jagannathan, Bharat; Golbeck, John (2009). "Breaking biological symmetry in membrane proteins: The asymmetrical orientation of PsaC on the pseudo-C2 symmetric Photosystem I core". Cell. Mol. Life Sci. 66 (7): 1257—1270. doi:10.1007/s00018-009-8673-x.
{{cite journal}}: Википедия:Обслуживание CS1 (множественные имена: authors list) (ссылка) - ↑ Jagannathan, Bharat; Golbeck, John (2009). "Understanding of the Binding Interface between PsaC and the PsaA/PsaB Heterodimer in Photosystem I". Biochemistry. 48: 5405—5416. doi:10.1021/bi900243f.
{{cite journal}}: Википедия:Обслуживание CS1 (множественные имена: authors list) (ссылка) - ↑ Ермаков, 2005, с. 157.
- ↑ PDB 3BQV.
- ↑ Frazão C, Sieker L, Sheldrick G, Lamzin V, LeGall J, Carrondo MA (April 1999). "Ab initio structure solution of a dimeric cytochrome c3 from Desulfovibrio gigas containing disulfide bridges". J. Biol. Inorg. Chem. 4 (2): 162—5. doi:10.1007/s007750050299. PMID 10499086.
{{cite journal}}: Википедия:Обслуживание CS1 (множественные имена: authors list) (ссылка) - ↑ Hope AB (January 2000). "Electron transfers amongst cytochrome f, plastocyanin and photosystem I: kinetics and mechanisms". Biochim. Biophys. Acta. 1456 (1): 5—26. doi:10.1016/S0005-2728(99)00101-2. PMID 10611452.
- ↑ Zhang L1, McSpadden B, Pakrasi HB, Whitmarsh J. (September 1992). "Copper-mediated regulation of cytochrome c553 and plastocyanin in the cyanobacterium Synechocystis 6803" (PDF). The journal of biological chemistry. 267 (27): 19054–19059. PMID 1326543.
{{cite journal}}: Википедия:Обслуживание CS1 (множественные имена: authors list) (ссылка) Википедия:Обслуживание CS1 (числовые имена: authors list) (ссылка) - ↑ 1 2 Rutherford A.W., Heathcote P. (1985). "Primary Photochemistry in Photosystem-I". Photosyn. Res. 6 (4): 295—316. doi:10.1007/BF00054105.
- ↑ 1 2 Zeiger, Eduardo; Taiz, Lincoln. Ch. 7: Topic 7.8: Photosystem I // Plant physiology. — 4th. — Sunderland, Mass : Sinauer Associates, 2006. — ISBN 0-87893-856-7.
- ↑ Shubin V.V., Karapetyan N.V., Krasnovsky A.A. (1986). "Molecular Arrangement of Pigment-Protein Complex of Photosystem I". Photosyn. Res. 9 (1–2): 3—12. doi:10.1007/BF00029726.
{{cite journal}}: Википедия:Обслуживание CS1 (множественные имена: authors list) (ссылка) - ↑ 1 2 Itoh, Shigeru, Msayo Iwaki (1989). "Vitamin K1 (Phylloquinone) Restores the Turnover of FeS centers of Ether-extracted Spinach PS I Particles". FEBS Lett. 243 (1): 47—52. doi:10.1016/0014-5793(89)81215-3.
{{cite journal}}: Википедия:Обслуживание CS1 (множественные имена: authors list) (ссылка) - ↑ Palace GP, Franke JE, Warden JT (May 1987). "Is phylloquinone an obligate electron carrier in photosystem I?". FEBS Lett. 215 (1): 58—62. doi:10.1016/0014-5793(87)80113-8. PMID 3552735.
{{cite journal}}: Википедия:Обслуживание CS1 (множественные имена: authors list) (ссылка) - ↑ 1 2 3 4 Vassiliev IR, Antonkine ML, Golbeck JH (October 2001). "Iron-sulfur clusters in type I reaction centers". Biochim. Biophys. Acta. 1507 (1–3): 139—60. doi:10.1016/S0005-2728(01)00197-9. PMID 11687212.
{{cite journal}}: Википедия:Обслуживание CS1 (множественные имена: authors list) (ссылка) - ↑ 1 2 Forti, Georgio, Paola Maria Giovanna Grubas (1985). "Two Sites of Interaction of Ferredoxin with thylakoids". FEBS Lett. 186 (2): 149—152. doi:10.1016/0014-5793(85)80698-0.
{{cite journal}}: Википедия:Обслуживание CS1 (множественные имена: authors list) (ссылка) - ↑ «The Photosynthetic Process» http://kentsimmons.uwinnipeg.ca/cm1504/lightreact.htm
- ↑ Robert Lucinski, Volkmar H.R. Schmid, Stefan Jansson, Frank Klimmek (2006). "Lhca5 interaction with plant photosystem I". FEBS letters. 580 (27): 6485—6488. doi:10.1016/j.febslet.2006.10.063.
{{cite journal}}: Википедия:Обслуживание CS1 (множественные имена: authors list) (ссылка) - ↑ 1 2 Lianwei Peng, Hiroshi Yamamoto, Toshiharu Shikanai (2011). "Structure and biogenesis of the chloroplast NAD(P)H dehydrogenase complex". Biochimica et Biophysica Acta (BBA). 1807 (8): 945—953. doi:10.1016/j.bbabio.2010.10.015.
{{cite journal}}: Википедия:Обслуживание CS1 (множественные имена: authors list) (ссылка) - ↑ Кренделева Т. Е., Кукарских Г. П., Тимофеев К. Н., Иванов Б. Н., Рубин А. Б. Ферредоксинзависимый циклический транспорт электронов в изолированных тилакоидах протекает с участием ферредоксин-НАДФ-редуктазы. Доклады академии наук, 2001. 379(5): с. 1-4.
- ↑ Коваленко И.Б., Устинин Д.М., Грачев Н.Е., Кренделева Т.Е., Кукарских Г.П., Тимофеев К.Н., Ризниченко Г.Ю., Грачев Е.А., Рубин А.Б. (2003). "Экспериментальное и теоретическое исследование процессов циклического электронного транспорта вокруг фотосистемы 1" (PDF). Биофизика. 48 (4): 656–665.
{{cite journal}}: Википедия:Обслуживание CS1 (множественные имена: authors list) (ссылка) - ↑ Cramer WA.; Zhang H.; Yan j.; Kurisu G.; Smith JL. (2006). "Transmembrane traffic in the cytochrome b6f complex". Annu Rev Biochem. 75: 769—90. doi:10.1146/annurev.biochem.75.103004.142756. PMID 16756511.
{{cite journal}}: Шаблон цитирования имеет пустые неизвестные параметры:|month=(справка)Википедия:Обслуживание CS1 (множественные имена: authors list) (ссылка) - ↑ Cramer WA.; Yan J.; Zhang H.; Kurisu G.; Smith JL. (2005). "Structure of the cytochrome b6f complex: new prosthetic groups, Q-space, and the 'hors d'oeuvres hypothesis' for assembly of the complex". Photosynth Res. 85 (1): 133—43. doi:10.1007/s11120-004-2149-5. PMID 15977064.
{{cite journal}}: Википедия:Обслуживание CS1 (множественные имена: authors list) (ссылка) - ↑ Masakazu Iwai, Kenji Takizawa, Ryutaro Tokutsu, Akira Okamuro, Yuichiro Takahashi & Jun Minagawa (22 April 2010). "Isolation of the elusive supercomplex that drives cyclic electron flow in photosynthesis". Nature. 464: 1210—1213. doi:10.1038/nature08885.
{{cite journal}}: Википедия:Обслуживание CS1 (множественные имена: authors list) (ссылка) - ↑ Hiroko Takahashi, Sophie Clowez, Francis-André Wollman, Olivier Vallon & Fabrice Rappaport (13 June 2013). "Cyclic electron flow is redox-controlled but independent of state transition". Nature Communications. 4. doi:10.1038/ncomms2954.
{{cite journal}}: Википедия:Обслуживание CS1 (множественные имена: authors list) (ссылка) - ↑ Lianwei Peng, Hideyuki Shimizu, Toshiharu Shikanai, (2008). "The Chloroplast NAD(P)H Dehydrogenase Complex Interacts with Photosystem I in Arabidopsis". J Biol Chem. 283 (50): 34873–34879. doi:10.1074/jbc.M803207200.
{{cite journal}}: Википедия:Обслуживание CS1 (лишняя пунктуация) (ссылка) Википедия:Обслуживание CS1 (множественные имена: authors list) (ссылка) Википедия:Обслуживание CS1 (не помеченный открытым DOI) (ссылка) - ↑ Yamori W, Sakata N, Suzuki Y, Shikanai T, Makino A. (2011). "Cyclic electron flow around photosystem I via chloroplast NAD(P)H dehydrogenase (NDH) complex performs a significant physiological role during photosynthesis and plant growth at low temperature in rice". Plant J. 68 (6): 966–76. doi:10.1111/j.1365-313X.2011.04747.x.
{{cite journal}}: Википедия:Обслуживание CS1 (множественные имена: authors list) (ссылка) - ↑ Chaddock, A.M.; Mant, A.; Karnauchov, I.; Brink, S.; Herrmann, R.G.; Klösgen, R.B.; Robinson, C. (1995). "A new type of signal peptide: central role of a twin-arginine motif in transfer signals for the delta pH-dependent thylakoidal protein translocase" (PDF). EMBO J. 14 (12): 2715—2722. PMC 398390. PMID 7796800.
- ↑ Kenneth Cline and Hiroki Mori (Aug 20, 2001). "Thylakoid ΔpH-dependent precursor proteins bind to a cpTatC–Hcf106 complex before Tha4-dependent transport". J Cell Biol. 154 (4): 719—730. doi:10.1083/jcb.200105149.
{{cite journal}}: Википедия:Обслуживание CS1 (дата и год) (ссылка) - ↑ McDonald AE, Ivanov AG, Bode R, Maxwell DP, Rodermel SR, Hüner NP (August 2011). "Flexibility in photosynthetic electron transport: the physiological role of plastoquinol terminal oxidase (PTOX)". Biochim. Biophys. Acta. 1807 (8): 954—67. doi:10.1016/j.bbabio.2010.10.024. PMID 21056542.
{{cite journal}}: Википедия:Обслуживание CS1 (множественные имена: authors list) (ссылка) - ↑ Ермаков, 2005, с. 123.
- ↑ Navassard V. Karapetyan, Alfred R. Holzwarth, Matthias Rögner (1999). "The photosystem I trimer of cyanobacteria: molecular organization, excitation dynamics and physiological significance". FEBS letters. 460 (3): 395—400. doi:10.1016/S0014-5793(99)01352-6.
{{cite journal}}: Википедия:Обслуживание CS1 (множественные имена: authors list) (ссылка) - ↑ Adam Ben-Shema, Felix Frolowb, Nathan Nelsona, (30 April 2004). "Evolution of photosystem I – from symmetry through pseudosymmetry to asymmetry". FEBS letters. 565 (3): 274—280. doi:10.1016/S0014-5793(04)00360-6.
{{cite journal}}: CS1 maint: year (ссылка) Википедия:Обслуживание CS1 (лишняя пунктуация) (ссылка) Википедия:Обслуживание CS1 (множественные имена: authors list) (ссылка) - ↑ Boudreau E, Takahashi Y, Lemieux C, Turmel M, Rochaix JD (1997). "The chloroplast ycf3 and ycf4 open reading frames of Chlamydomonas reinhardtii are required for the accumulation of the photosystem I complex". EMBO J. 16 (20): 6095—104. doi:10.1093/emboj/16.20.6095. PMC 1326293. PMID 9321389.
{{cite journal}}: Википедия:Обслуживание CS1 (множественные имена: authors list) (ссылка) - ↑ 1 2 3 4 5 Lockau, Wolfgang, Wolfgang Nitschke (1993). "Photosystem I and its Bacterial Counterparts". Physiologia Plantarum. 88 (2): 372—381. doi:10.1111/j.1399-3054.1993.tb05512.x.
{{cite journal}}: Википедия:Обслуживание CS1 (множественные имена: authors list) (ссылка)
Литература
- Зитте П. и др. Ботаника / Под ред. В. В. Чуба. — 35-е изд. — М.: Академия, 2008. — Т. 2. Физиология растений. — 495 с.
- Медведев С. С. Физиология растений. — СПб.: БХВ-Петербург, 2013. — 335 с.
- Физиология растений / Под ред. И. П. Ермакова. — М.: Академия, 2005. — 634 с.
- Хелдт Г. В. Биохимия растений. — М.: БИНОМ. Лаборатория знаний, 2011. — 471 с.
Ссылки
- Информационная система «Фотосинтетическая мембрана»
- «Циклический и нециклический поток электронов.» в онлайн энциклопедии Физиология растений
Эта статья входит в число хороших статей русскоязычного раздела Википедии. |