Фосфоенолпируваткарбоксикиназа: различия между версиями
| [отпатрулированная версия] | [отпатрулированная версия] |
Djem92 (обсуждение | вклад) |
Djem92 (обсуждение | вклад) |
||
| Строка 49: | Строка 49: | ||
== Регуляция == |
== Регуляция == |
||
=== У человека === |
=== У человека === |
||
Усиление [[Экспрессия генов|экспрессии]] и активации PEPCK-С (цитозольная форма) происходит под действием многих факторов. Транскрипция гена PEPCK-C стимулируется [[глюкагон]]ом, [[Глюкокортикоиды|глюкокортикоидами]], [[Ретиноевая кислота|ретиноевой кислотой]] и аденозин-3',5'-монофосфатом (цАМФ), а [[инсулин]] (посредством [[ |
Усиление [[Экспрессия генов|экспрессии]] и активации PEPCK-С (цитозольная форма) происходит под действием многих факторов. Транскрипция гена PEPCK-C стимулируется [[глюкагон]]ом, [[Глюкокортикоиды|глюкокортикоидами]], [[Ретиноевая кислота|ретиноевой кислотой]] и аденозин-3',5'-монофосфатом (цАМФ), а [[инсулин]] (посредством [[SREBP1]]c) её подавляет<ref name="Obrien">{{cite journal | vauthors = O'Brien RM, Lucas PC, Forest CD, Magnuson MA, Granner DK | title = Identification of a sequence in the PEPCK gene that mediates a negative effect of insulin on transcription | journal = Science | volume = 249 | issue = 4968 | pages = 533–7 | date = August 1990 | pmid = 2166335 | doi = 10.1126/science.2166335 | bibcode = 1990Sci...249..533O }}</ref>. Из этих факторов инсулин, гормон, дефицит которого наблюдается при [[Сахарный диабет 1-го типа|сахарном диабете 1-го типа]], считается доминирующим, поскольку он подавляет транскрипцию многих стимулирующих элементов<ref name="Obrien"/>. Активность PEPCK также ингибируется [[Сульфат гидразиния|гидразинсульфат]]ом, и это ингибирование снижает скорость [[глюконеогенез]]а<ref name="Mazzio">{{cite journal | vauthors = Mazzio E, Soliman KF | title = The role of glycolysis and gluconeogenesis in the cytoprotection of neuroblastoma cells against 1-methyl 4-phenylpyridinium ion toxicity | journal = Neurotoxicology | volume = 24 | issue = 1 | pages = 137–47 | date = January 2003 | pmid = 12564389 | doi = 10.1016/S0161-813X(02)00110-9 }}</ref>. |
||
При длительном [[ацидоз]]е в клетках щёточной каймы проксимальных канальцев почек повышается активность PEPCK-C, что приводит к выделению большего количества [[аммиак]]а NH<sub>3</sub> и, соответственно, к образованию большего количества гидрокарбонат ионов HCO<sub>3</sub><sup>-</sup><ref name=boron858>{{cite book |author=Walter F. Boron |title=Medical Physiology: A Cellular And Molecular Approach |publisher=Elsevier/Saunders |page=858 |isbn=978-1-4160-2328-9 |year=2005 }}</ref>. |
При длительном [[ацидоз]]е в клетках щёточной каймы проксимальных канальцев почек повышается активность PEPCK-C, что приводит к выделению большего количества [[аммиак]]а NH<sub>3</sub> и, соответственно, к образованию большего количества гидрокарбонат ионов HCO<sub>3</sub><sup>-</sup><ref name=boron858>{{cite book |author=Walter F. Boron |title=Medical Physiology: A Cellular And Molecular Approach |publisher=Elsevier/Saunders |page=858 |isbn=978-1-4160-2328-9 |year=2005 }}</ref>. |
||
Версия от 20:09, 25 ноября 2023
| Фосфоенолпируваткарбоксикиназа | |
|---|---|
 PDB rendering based on 1khb. | |
| Идентификаторы | |
| Символ | PEPCK |
| Pfam | PF00821 |
| InterPro | IPR008209 |
| PROSITE | PDOC00421 |
| SCOP | 1khf |
| SUPERFAMILY | 1khf |
| Доступные структуры белков | |
| Pfam | структуры |
| PDB | RCSB PDB; PDBe; PDBj |
| PDBsum | 3D-модель |
Фосфоенолпируваткарбоксикиназа, также ФЕП-карбоксикиназа (англ. Phosphoenolpyruvate carboxykinase, сокр. PEPCK) — фермент (КФ 4.1.1.32) из семейства декарбоксилаз (класс лиазы), катализирующий реакцию декарбоксилирования-фосфорилирования молекул оксалоацетата с образованием фосфоенолпирувата, углекислого газа[1][2][3] и GDP. В качестве источника неорганического фосфата фермент использует молекулы GTP, а кофактором могут служить ионы Mn2+. Схема реакции:
- Оксалоацетат + GTP ↔ Фосфоенолпируват + СО2 + GDP.
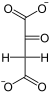 + GTP
+ GTP  + CO2 + GDP
+ CO2 + GDP



Данная реакция относится к так называемым «обходным реакциям» гликолиза. ФЕП-карбоксикиназа играет ключевую роль в процессе глюконеогенеза. Существует две изоформы фермента, цитозольная и митохондриальная. Ген, кодирующий митохондриальную форму фермента — PCK2 (также PEPCK-M) у человека локализован на длинном плече (q-плече) 14-й хромосомы. Ген цитозольной формы — PCK1 или PEPCK-C располагается на длинном плече (q-плече) 20-й хромосомы.
ФЕП-карбоксикиназа (митохондриальная) состоит из 640 аминокислотных остатков и имеет молекулярную массу — 70699 Да. Цитозольная форма содержит 622 аминокислоты и имеет молекулярную массу — 69195 Да.
Структура

У человека присутствуют два изофермента ФЕП-карбоксикиназы: цитозольный (SwissProt P35558) и митохондриальный (SwissProt Q16822), которые имеют 63,4 % идентичности. Цитозольная форма играет важную роль в глюконеогенезе. Однако известен механизм переноса фосфоенолпирувата (PEP) из митохондрий в цитозоль с помощью специфических мембранных транспортных белков[4][5][6][7][8]. В переносе PEP через внутреннюю митохондриальную мембрану участвует митохондриальный трикарбоксилатный транспортный белок и в меньшей степени переносчик адениновых нуклеотидов. Выдвигается также предположение о возможности существования транспортёра PEP/пирувата[9].
Рентгеновские структуры PEPCK дают представление о структуре и механизме каталитической активности PEPCK. Митохондриальная изоформа PEPCK из куриной печени в комплексе с ионами Mn2+, Mn2+-фосфоенолпируватом (PEP) и Mn2+-GDP даёт информацию о её структуре и о том, как этот фермент катализирует реакции[10]. Delbaere et al. (2004 год) обнаружили PEPCK в клетках E. coli и выявили активный сайт, расположенный между C-концевым доменом и N-концевым доменом. Было обнаружено, что активный центр фермента закрывается при вращении этих доменов[11].
В процессе действия ФЕП-карбоксикиназы происходит перенос фосфорильных групп, чему, вероятно, способствует заслонённая конформация фосфорильных групп при связывании молекул ATP с ферментом[11].
Поскольку заслонённая конформация обладает высокой энергией, перенос фосфорильных групп имеет пониженную энергию активации, что означает эти группы будут переноситься с большей легкостью. Этот перенос, вероятно, происходит по механизму, аналогичному нуклеофильному замещению SN2[11].
У различных видов
Транскрипция гена PEPCK происходит у многих видов, и аминокислотная последовательность фермента у каждого вида своя.
Например, его структура и специфичность различаются у человека, кишечной палочки (E. coli) и паразита Trypanosoma cruzi[12].
Механизм катализа
Поскольку ФЕП-карбоксикиназа действует на стыке гликолиза и цикла Кребса, она вызывает декарбоксилирование молекулы оксалоацетата (C4) с образованием молекулы PEP (C3). В качестве первого обязательного этапа глюконеогенеза PEPCK декарбоксилирует и фосфорилирует оксалоацетат для его преобразования в PEP, в присутствии молекул GTP. При переносе фосфата в результате реакции образуется молекула GDP[10]. В активном центре фермента имеется остаток аргинина и ионы двухвалентного марганца Mn2+. Кислород кетогруппы (C=O) оксалоацетата образует комплекс между аргинином и Mn2+ и, таким образом, позволяет отщепить карбоксильную группу на более дальнем конце с образованием двойной связи. Затем фосфат переносится от GTP к кислороду. В то же время ионы Mn2+ и аргинин также обеспечивают комплексообразование GTP в ферменте[13].
Когда пируваткиназа – фермент, который обычно катализирует реакцию, превращающая PEP в пируват — нокаутируется у мутантов Bacillus subtilis, PEPCK участвует в одной из замещающих анаплеротических реакций, действуя в направлении, противоположному нормального катализа, превращая PEP в оксалоацетат[14]. Хотя эта реакция и возможна, кинетика настолько неблагоприятна, что мутанты растут очень медленно или вообще не растут[14].
Регуляция
У человека
Усиление экспрессии и активации PEPCK-С (цитозольная форма) происходит под действием многих факторов. Транскрипция гена PEPCK-C стимулируется глюкагоном, глюкокортикоидами, ретиноевой кислотой и аденозин-3',5'-монофосфатом (цАМФ), а инсулин (посредством SREBP1c) её подавляет[15]. Из этих факторов инсулин, гормон, дефицит которого наблюдается при сахарном диабете 1-го типа, считается доминирующим, поскольку он подавляет транскрипцию многих стимулирующих элементов[15]. Активность PEPCK также ингибируется гидразинсульфатом, и это ингибирование снижает скорость глюконеогенеза[16].
При длительном ацидозе в клетках щёточной каймы проксимальных канальцев почек повышается активность PEPCK-C, что приводит к выделению большего количества аммиака NH3 и, соответственно, к образованию большего количества гидрокарбонат ионов HCO3-[17].
ГТФ-специфическая активность PEPCK наиболее высока при наличии двухвалентных ионов — Mn2+ и Mg2+[18]. Кроме того, гиперреактивный цистеин (Cys307) фермента участвует в связывании Mn2+ с активным сайтом[10].
У растений
Как уже говорилось ранее, при поливе растений хлористым аммонием с низким pH концентрация ФЭП-карбоксикиназы увеличивалась, хотя при высоком pH такого эффекта не наблюдалось
Функции
Глюконеогенез
PEPCK-C (цитозольная форма) катализирует необратимый этап глюконеогенеза — процесс синтеза глюкозы. Поэтому считается, что этот фермент играет важную роль в гомеостазе глюкозы, что подтверждается на примере лабораторных мышей, заболевших сахарным диабетом 2-го типа в результате сверхэкспрессии гена PEPCK-C[19].
Роль PEPCK-C в глюконеогенезе может быть опосредована циклом лимонной кислоты, активность которого, как было установлено, напрямую зависит от количества PEPCK-C[20].
Уровни PEPCK-C сами по себе не сильно коррелировали с глюконеогенезом в печени мышей, как предполагали предыдущие исследования[20]. В то время как печень мыши почти исключительно экспрессирует PEPCK-C, у людей в равной степени присутствует митохондриальный изофермент (PEPCK-M). PEPCK-M сам по себе обладает глюконеогенным потенциалом[2]. Следовательно, роль PEPCK-C и PEPCK-M в глюконеогенезе может быть более сложной и включать больше факторов, чем считалось ранее.
У растений
ФЕП-карбоксикиназа (КФ 4.1.1.49) — один из трёх ферментов декарбоксилирования, используемых в механизмах концентрирования неорганического углерода в растениях с C4-фотосинтезом и CAM. Другие — NADP-зависимая малатдегидрогеназа и NAD-зависимая малатдегидрогеназа (малик-фермент)[21][22]. При фиксации углерода в С4-растениях углекислый газ сначала фиксируется путём присоединения к фосфоенолпирувату с образованием оксалоацетата в мезофилле. В растениях с С4-фотосинтезом оксалоацетат затем преобразуется ферментом PEPCK в аспартат, который поступает в проводящий пучок. В клетках проводящего пучка аспартат снова превращается в оксалоацетат. PEPCK декарбоксилирует оксалоацетат оболочки проводящего пучка, выделяя углекислый газ, который затем фиксируется ферментом Rubisco. На каждую молекулу углекислого газа, произведенную PEPCK, расходуется молекула ATP.
PEPCK активна в растениях, которые подвергаются фиксации углерода путём C4-фотосинтеза, где её действие локализовано в цитозоле, в отличие от млекопитающих, у которых было обнаружено, что PEPCK активна также в митохондриях[23].
Хотя ФЕП-карбоксикиназа обнаружена во многих частях растений, её активность была отмечена только в определённых типах клеток, в том числе в участках флоэмы[24].
Также было обнаружено, что в огурце (Cucumis sativus L.) концентрация PEPCK повышается под действием многочисленных эффектов, которые, как известно, снижают значение pH клеток растений, хотя эти эффекты специфичны для части растения[24].
Уровни PEPCK повышались в корнях и стеблях, когда растения поливали хлоридом аммония при низком значении pH (но не при высоком pH) или масляной кислотой. Однако уровни PEPCK в листьях в этих условиях не увеличивались.
В листьях, при 5%-ном содержание CO2 в атмосфере, происходит увеличение количества ФЕП-карбоксикиназы[24].
У бактерий
В попытке изучить роль ФЕП-карбоксикиназы исследователи вызвали сверхэкспрессию гена PEPCK в бактериях E. coli с помощью рекомбинантной ДНК[18].
Было показано, что PEPCK Mycobacterium tuberculosis запускает иммунную систему у мышей, за счёт повышения активности цитокинов[25].
В результате было установлено, что PEPCK может быть подходящим компонентом для разработки эффективной субъединичной вакцины против туберкулёза[25].
Клиническое значение
PEPCK до недавнего времени не рассматривался в исследованиях рака. Было показано, что в образцах опухолей человека и линиях раковых клеток человека (клетки рака молочной железы, толстой кишки и лёгких) PEPCK-M, а не PEPCK-C, экспрессировался на достаточных уровнях, чтобы играть соответствующую метаболическую роль[1][26]. Таким образом, PEPCK-M может играть роль в раковых клетках, особенно при ограничении питательных веществ или других стрессовых условиях.
Примечания
- ↑ 1 2 Méndez-Lucas A, Hyroššová P, Novellasdemunt L, Viñals F, Perales JC (August 2014). "Mitochondrial phosphoenolpyruvate carboxykinase (PEPCK-M) is a pro-survival, endoplasmic reticulum (ER) stress response gene involved in tumor cell adaptation to nutrient availability". The Journal of Biological Chemistry. 289 (32): 22090—102. doi:10.1074/jbc.M114.566927. PMC 4139223. PMID 24973213.
- ↑ 1 2 Méndez-Lucas A, Duarte JA, Sunny NE, Satapati S, He T, Fu X, et al. (July 2013). "PEPCK-M expression in mouse liver potentiates, not replaces, PEPCK-C mediated gluconeogenesis". Journal of Hepatology. 59 (1): 105—13. doi:10.1016/j.jhep.2013.02.020. PMC 3910155. PMID 23466304.
- ↑ Chakravarty K, Cassuto H, Reshef L, Hanson RW (2005). "Factors that control the tissue-specific transcription of the gene for phosphoenolpyruvate carboxykinase-C". Critical Reviews in Biochemistry and Molecular Biology. 40 (3): 129—54. doi:10.1080/10409230590935479. PMID 15917397. S2CID 633399.
- ↑ Robinson BH (May 1971). "Transport of phosphoenolpyruvate by the tricarboxylate transporting system in mammalian mitochondria". FEBS Letters. 14 (5): 309—312. doi:10.1016/0014-5793(71)80287-9. PMID 11945784. S2CID 9617975.
- ↑ Söling HD, Walter U, Sauer H, Kleineke J (December 1971). "Effects of synthetic analogues of phosphoenolpyruvate on muscle and liver pyruvate kinase, muscle enolase, liver phosphoenolpyruvate carboxykinase and on the intra-/extra-mitochondrial tricarboxylic acid carrier transport system". FEBS Letters. 19 (2): 139—143. doi:10.1016/0014-5793(71)80498-2. PMID 11946196. S2CID 40637963.
- ↑ Kleineke J, Sauer H, Söling HD (January 1973). "On the specificity of the tricarboxylate carrier system in rat liver mitochondria". FEBS Letters. 29 (2): 82—6. doi:10.1016/0014-5793(73)80531-9. PMID 4719206. S2CID 30730789.
- ↑ Shug AL, Shrago E (July 1973). "Inhibition of phosphoenolpyruvate transport via the tricarboxylate and adenine nucleotide carrier systems of rat liver mitochondria". Biochemical and Biophysical Research Communications. 53 (2): 659—65. doi:10.1016/0006-291X(73)90712-2. PMID 4716993.
- ↑ Sul HS, Shrago E, Shug AL (January 1976). "Relationship of phosphoenolpyruvate transport, acyl coenzyme A inhibition of adenine nucleotide translocase and calcium ion efflux in guinea pig heart mitochondria". Archives of Biochemistry and Biophysics. 172 (1): 230—7. doi:10.1016/0003-9861(76)90071-0. PMID 1252077.
- ↑ Satrústegui J, Pardo B, Del Arco A (January 2007). "Mitochondrial transporters as novel targets for intracellular calcium signaling". Physiological Reviews. 87 (1): 29—67. doi:10.1152/physrev.00005.2006. PMID 17237342.
- ↑ 1 2 3 Holyoak T, Sullivan SM, Nowak T (July 2006). "Structural insights into the mechanism of PEPCK catalysis". Biochemistry. 45 (27): 8254—63. doi:10.1021/bi060269g. PMID 16819824.
- ↑ 1 2 3 Delbaere LT, Sudom AM, Prasad L, Leduc Y, Goldie H (March 2004). "Structure/function studies of phosphoryl transfer by phosphoenolpyruvate carboxykinase". Biochimica et Biophysica Acta (BBA) - Proteins and Proteomics. 1697 (1—2): 271—8. doi:10.1016/j.bbapap.2003.11.030. PMID 15023367.
- ↑ Trapani S, Linss J, Goldenberg S, Fischer H, Craievich AF, Oliva G (November 2001). "Crystal structure of the dimeric phosphoenolpyruvate carboxykinase (PEPCK) from Trypanosoma cruzi at 2 A resolution". Journal of Molecular Biology. 313 (5): 1059—72. doi:10.1006/jmbi.2001.5093. PMID 11700062.
- ↑ L. W. Tari: Structure and Mechanism of Phosphoenolpyruvate Carboxykinase. In: Journal of Biological Chemistry. 272, S. 8105–8108, doi:10.1074/jbc.272.13.8105.
- ↑ 1 2 Zamboni N, Maaheimo H, Szyperski T, Hohmann HP, Sauer U (October 2004). "The phosphoenolpyruvate carboxykinase also catalyzes C3 carboxylation at the interface of glycolysis and the TCA cycle of Bacillus subtilis". Metabolic Engineering. 6 (4): 277—84. doi:10.1016/j.ymben.2004.03.001. PMID 15491857.
- ↑ 1 2 O'Brien RM, Lucas PC, Forest CD, Magnuson MA, Granner DK (August 1990). "Identification of a sequence in the PEPCK gene that mediates a negative effect of insulin on transcription". Science. 249 (4968): 533—7. Bibcode:1990Sci...249..533O. doi:10.1126/science.2166335. PMID 2166335.
- ↑ Mazzio E, Soliman KF (January 2003). "The role of glycolysis and gluconeogenesis in the cytoprotection of neuroblastoma cells against 1-methyl 4-phenylpyridinium ion toxicity". Neurotoxicology. 24 (1): 137—47. doi:10.1016/S0161-813X(02)00110-9. PMID 12564389.
- ↑ Walter F. Boron. Medical Physiology: A Cellular And Molecular Approach. — Elsevier/Saunders, 2005. — P. 858. — ISBN 978-1-4160-2328-9.
- ↑ 1 2 Aich S, Imabayashi F, Delbaere LT (October 2003). "Expression, purification, and characterization of a bacterial GTP-dependent PEP carboxykinase". Protein Expression and Purification. 31 (2): 298—304. doi:10.1016/S1046-5928(03)00189-X. PMID 14550651.
- ↑ Vanderbilt Medical Center. "Granner Lab, PEPCK Research." 2001. Online. Internet. Accessed 10:46PM, 4/13/07. www.mc.vanderbilt.edu/root/vumc.php?site=granner&doc=119
- ↑ 1 2 Burgess SC, He T, Yan Z, Lindner J, Sherry AD, Malloy CR, et al. (April 2007). "Cytosolic phosphoenolpyruvate carboxykinase does not solely control the rate of hepatic gluconeogenesis in the intact mouse liver". Cell Metabolism. 5 (4): 313—20. doi:10.1016/j.cmet.2007.03.004. PMC 2680089. PMID 17403375.
- ↑ Kanai R, Edwards, GE. The Biochemistry of C4 Photosynthesis // C4 Plant Biology. — Elsevier, 1998. — P. 49–87. — ISBN 978-0-08-052839-7.
- ↑ Christopher JT, Holtum J (September 1996). "Patterns of Carbon Partitioning in Leaves of Crassulacean Acid Metabolism Species during Deacidification". Plant Physiology. 112 (1): 393—399. doi:10.1104/pp.112.1.393. PMC 157961. PMID 12226397.
- ↑ Voznesenskaya EV, Franceschi VR, Chuong SD, Edwards GE (July 2006). "Functional characterization of phosphoenolpyruvate carboxykinase-type C4 leaf anatomy: immuno-, cytochemical and ultrastructural analyses". Annals of Botany. 98 (1): 77—91. doi:10.1093/aob/mcl096. PMC 2803547. PMID 16704997.
- ↑ 1 2 3 Chen ZH, Walker RP, Técsi LI, Lea PJ, Leegood RC (May 2004). "Phosphoenolpyruvate carboxykinase in cucumber plants is increased both by ammonium and by acidification, and is present in the phloem". Planta. 219 (1): 48—58. doi:10.1007/s00425-004-1220-y. PMID 14991407. S2CID 23800457.
- ↑ 1 2 Liu K, Ba X, Yu J, Li J, Wei Q, Han G, et al. (August 2006). "The phosphoenolpyruvate carboxykinase of Mycobacterium tuberculosis induces strong cell-mediated immune responses in mice". Molecular and Cellular Biochemistry. 288 (1—2): 65—71. doi:10.1007/s11010-006-9119-5. PMID 16691317. S2CID 36284611.
- ↑ Leithner K, Hrzenjak A, Trötzmüller M, Moustafa T, Köfeler HC, Wohlkoenig C, et al. (February 2015). "PCK2 activation mediates an adaptive response to glucose depletion in lung cancer". Oncogene. 34 (8): 1044—50. doi:10.1038/onc.2014.47. PMID 24632615. S2CID 11902696.
| Гликолиз | |||||||||
|---|---|---|---|---|---|---|---|---|---|
| Только глюконеогенез |
| ||||||||
| Регуляция | |||||||||