Белки, заякоренные липидами

Белки́, зая́коренные липи́дами, или липидосвя́занные белки́ (англ. Lipid-anchored proteins, lipid-linked proteins) — белки клеточной мембраны, ковалентно связанные с липидами клеточной мембраны. Эти липиды вставлены в мембрану бок о бок с хвостами жирных кислот. Белки, заякоренные липидами, могут находиться с любой стороны клеточной мембраны. Таким образом, липид служит своего рода якорем, закрепляющим белок вблизи клеточной мембраны[1][2].
Липидные группы, связанные с белками, могут принимать участие в белок-белковых взаимодействиях и влиять на работу белков, которые к ним прикреплены[2][3]. Например, липидные группы могут играть важную роль в увеличении гидрофобности молекулы. Это обеспечивает способность заякоренных белков взаимодействовать с клеточной мембраной и белковыми доменами[4].
Существует три типа белков, заякоренных липидами:
- пренилированные белки;
- белки, ацилированные жирными кислотами;
- белки, связанные с гликозилфосфатидилинозитолом (GPI)[2].
К белку могут быть ковалентно присоединены несколько липидных групп, однако место, которым белок связан с липидом, зависит и от липида, и от самого белка[2].
Пренилированные белки
[править | править код]

Как и следует из названия, пренилированные белки ковалентно связаны с гидрофобными полимерами изопрена (то есть разветвлённого пятиуглеродного углеводорода[5]) через аминокислотные остатки цистеина[2][3]. Изопреноидные группы, обычно фарнезил (15 атомов углерода) и геранилгеранил[англ.] (20 атомов углерода), прикрепляются к белку при помощи тиоэфирных связей с остатками цистеина, расположенными рядом с С-концом белка[3][4]. Пренилирование белков облегчает их взаимодействие с клеточной мембраной[1].

Мотив пренилирования, известный как CAAX-бокс — наиболее популярный сайт пренилирования белков, то есть сайт, к которому ковалентно прикрепляются фарнезил или геранилгеранил[2][3]. В CAAX-боксе С — это пренилируемый остаток цистеина, А — алифатический аминокислотный остаток и Х определяет, какой тип пренилирования будет иметь место. Если Х — это Ala, Met, Ser или Gln, то белок будет фарнезилирован при помощи фермента фарнезилтрансферазы[англ.], а если Х — это Leu, то белок будет геранилгеранилирован ферментом геранилгеранилтрансферазой I[англ.][3][4]. Эти ферменты схожи и оба состоят из двух субъединиц[6].
Пренилированные белки очень важны для роста эукариотической клетки, дифференцировки и морфологии[6]. Более того, пренилирование — это обратимая посттрансляционная модификация. Динамическое взаимодействие пренилированных белков с клеточной мембраной необходимо для их участия в передаче сигнала и часто расстроено при таких заболеваниях, как рак[7] . Например, Ras — это белок, подвергающийся пренилированию фарнезилтрансферазой, и когда он находится в активном состоянии, он может включать гены, участвующие в клеточном росте и дифференцировке. Поэтому чрезмерная активность Ras может привести к раку[8]. Изучение механизма действия пренилированных белков важно для разработки противораковых препаратов[9]. К пренилированным белкам также относятся члены белковых семейств Rab[англ.] и Rho, а также ламины[6].
Некоторые пренильные группы, участвующие в метаболическом пути HGM-КоА редуктазы[1], — геранилгераниол[англ.], фарнезол и долихол — в связанном с пирофосфатом виде участвуют в реакциях конденсации, ускоряемых такими ферментами, как пренилтрансфераза[англ.], и в итоге образуют циклы с образованием холестерина[2].
Белки, ацилированные жирными кислотами
[править | править код]Белки, ацилированные жирными кислотами, — это белки, которые подверглись посттрансляционным модификациям и стали ковалентно связанными с жирными кислотами по некоторым аминокислотным остаткам[10][11]. К числу жирных кислот, наиболее часто связанных с белками, относят насыщенную 14-углеродную миристиновую кислоту и 16-углеродную пальмитиновую кислоту. Белки могут быть связаны как с одной из этих жирных кислот, так и с двумя[10].
N-миристоилирование
[править | править код]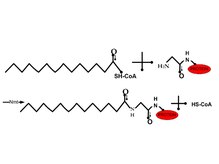
N-миристоилирование (то есть присоединение миристиновой кислоты) — это, как правило, необратимая белковая модификация, которая обычно происходит во время синтеза белка[10][12] и представляет собой присоединение миристиновой кислоты к α-аминогруппы концевого остатка глицина посредством пептидной связи[2][11]. Эта реакция ускоряется ферментом N-миристоилтрансферазой[англ.]. Белки, подвергающиеся N-миристоилированию, начинаются с Met-Gly и имеют серин или треонин в позиции 5[10]. Миристоилированные белки участвуют в передаче сигнала, белок-белковых взаимодействиях и механизмах, которые регулируют нацеливание и функционирование других белков[12]. Например, миристоилирование белка Bid[англ.] важно для регуляции апоптоза: миристоилированный Bid направляется к митохондриям и вызывает выход оттуда цитохрома с, что в конечном счёте приводит к апоптозу. Другие миристоилированные белки, участвующие в регуляции апоптоза, — актин и гельзолин[англ.][13].
S-пальмитоилирование
[править | править код]
S-пальмитоилирование (то есть присоединение пальмитиновой кислоты) — это обратимая белковая модификация, при которой пальмитиновая кислота присоединяется к специфическому остатку цистеина посредством тиоэфирной связи[2][10]. Когда к пальмитоилированным белкам присоединяются жирные кислоты средней и большой длины, может использоваться термин S-ацилирование. Для пальмитоилирования не было определено консенсусной последовательности[10]. Пальмитолированные белки в основном встречаются на цитоплазматической стороне клеточной мембраны, где они участвуют в передаче сигнала[2]. Пальмитоильная группа может быть удалена ферментами пальмитоилтиоэстеразами. Предполагается, что обратимое пальмитоилирование может регулировать взаимодействие белка с клеточной мембраной и таким образом принимать участие в передаче сигнала. Кроме того, оно может использоваться для регуляции внутриклеточной локализации белка, стабильности и кругооборота[14]. Например, в синапсе пальмитоилирование белков играет ключевую роль в передаче сигнала, регулируя кластеризацию белков. Когда белок PSD-95[англ.] пальмитоилирован, он связан с мембраной и может связываться и кластеризовать ионные каналы на постсинаптической мембране. Таким образом, пальмитоилирование может играть роль в регуляции высвобождения нейромедиаторов[15].
GPI-белки
[править | править код]
GPI-белки присоединяются к комплексу GPI через пептидную связь С-концевой карбоксильной группы белка[16]. GPI-белок состоит из нескольких связанных компонентов: фосфоэтаноламина[англ.], линейного тетрасахарида (состоящего из трёх остатков маннозы и одного глюкозаминила) и фосфатидилинозитола[17]. Фосфатидилинозитол связан гликозидной связью с не-N-ацетилированным глюкозамином тетрасахарида. Между маннозой на нередуцирующем конце тетрасахарида и фосфоэтаноламином образуется фосфодиэфирная связь. Фосфоэтаноламин далее связан с С-концом соответствующего белка пептидной связью[2]. Прикрепление белка к комплексу GPI опосредуется ферментным комплексом GPI-трансамидаза[17]. Жирные кислоты фосфатидилинозитола вставлены в мембрану и заякоривают белок[16]. GPI-белки располагаются только с внешней стороны клеточной мембраны[2].
Остатки сахаров в тетрасахариде и остатки жирных кислот фосфатидилинозитола варьируют от белка к белку[2]. Благодаря этому разнообразию GPI-белки могут выполнять самые разнообразные функции: выступая как гидролитические ферменты, адгезионные молекулы, рецепторы, ингибиторы протеаз[18]. Более того, GPI-белки играют важные роли в эмбриогенезе, развитии, нейрогенезе, работе иммунной системы и оплодотворении[16]. Например, GPI-белок IZUMO1R/JUNO, названный в честь древнеримской богини плодородия и расположенный на мембране яйцеклетки, необходим для слияния яйцеклетки и сперматозоида. Без этого белка яйцеклетка и сперматозоид не могут слиться, поэтому его исчезнвение после оплодотворения может быть одним из механизмов, защищающих от полиспермии[19]. Другие GPI-белки участвуют в ассоциации мембранных микродоменов, временной гомодимеризации и апикальной сортировке у поляризованных клеток[16].
Примечания
[править | править код]- ↑ 1 2 3 Gerald Karp. Cell and Molecular Biology: Concepts and Experiments (англ.). — John Wiley and Sons, 2009. — P. 128—. — ISBN 978-0-470-48337-4.
- ↑ 1 2 3 4 5 6 7 8 9 10 11 12 13 Voet Judith G., Pratt Charlotte W. Fundamentals of Biochemistry: Life at the Molecular Level (англ.). — 4th. — John Wiley & Sons, Inc., 2013. — P. 263. — ISBN 978-0470-54784-7.
- ↑ 1 2 3 4 5 Casey P. J., Seabra M. C. Protein prenyltransferases. (англ.) // The Journal of biological chemistry. — 1996. — Vol. 271, no. 10. — P. 5289—5292. — PMID 8621375.
- ↑ 1 2 3 Novelli Giuseppe, D’Apice Maria Rosaria. Protein farnesylation and disease // Journal of Inherited Metabolic Disease. — 2012. — 4 февраля (т. 35, № 5). — С. 917—926. — ISSN 0141-8955. — doi:10.1007/s10545-011-9445-y.
- ↑ isoprene Miller-Keane Encyclopedia and Dictionary of Medicine, Nursing, and Allied Health, Seventh Ed (2003). Дата обращения: 28 ноября 2015. Архивировано 8 декабря 2015 года.
- ↑ 1 2 3 Lane K. T., Beese L. S. Thematic review series: lipid posttranslational modifications. Structural biology of protein farnesyltransferase and geranylgeranyltransferase type I. (англ.) // Journal of lipid research. — 2006. — Vol. 47, no. 4. — P. 681—699. — doi:10.1194/jlr.R600002-JLR200. — PMID 16477080.
- ↑ Stein V., Kubala M. H., Steen J., Grimmond S. M., Alexandrov K. Towards the systematic mapping and engineering of the protein prenylation machinery in Saccharomyces cerevisiae. (англ.) // Public Library of Science ONE. — 2015. — Vol. 10, no. 3. — P. e0120716. — doi:10.1371/journal.pone.0120716. — PMID 25768003.
- ↑ Goodsell D. S. The molecular perspective: the ras oncogene. (англ.) // The oncologist. — 1999. — Vol. 4, no. 3. — P. 263—264. — PMID 10394594.
- ↑ Reuter C. W., Morgan M. A., Bergmann L. Targeting the Ras signaling pathway: a rational, mechanism-based treatment for hematologic malignancies? (англ.) // Blood. — 2000. — Vol. 96, no. 5. — P. 1655—1669. — PMID 10961860.
- ↑ 1 2 3 4 5 6 Resh Marilyn D. Trafficking and signaling by fatty-acylated and prenylated proteins // Nature Chemical Biology. — 2006. — Ноябрь (т. 2, № 11). — С. 584—590. — ISSN 1552-4450. — doi:10.1038/nchembio834.
- ↑ 1 2 Wilson J. P., Raghavan A. S., Yang Y. Y., Charron G., Hang H. C. Proteomic analysis of fatty-acylated proteins in mammalian cells with chemical reporters reveals S-acylation of histone H3 variants. (англ.) // Molecular & cellular proteomics : MCP. — 2011. — Vol. 10, no. 3. — P. 110—001198. — doi:10.1074/mcp.M110.001198. — PMID 21076176.
- ↑ 1 2 Farazi T. A., Waksman G., Gordon J. I. The biology and enzymology of protein N-myristoylation. (англ.) // The Journal of biological chemistry. — 2001. — Vol. 276, no. 43. — P. 39501—39504. — doi:10.1074/jbc.R100042200. — PMID 11527981.
- ↑ Martin Dale D.O., Beauchamp Erwan, Berthiaume Luc G. Post-translational myristoylation: Fat matters in cellular life and death // Biochimie. — 2011. — Январь (т. 93, № 1). — С. 18—31. — ISSN 0300-9084. — doi:10.1016/j.biochi.2010.10.018.
- ↑ Aicart-Ramos Clara, Valero Ruth Ana, Rodriguez-Crespo Ignacio. Protein palmitoylation and subcellular trafficking // Biochimica et Biophysica Acta (BBA) - Biomembranes. — 2011. — Декабрь (т. 1808, № 12). — С. 2981—2994. — ISSN 0005-2736. — doi:10.1016/j.bbamem.2011.07.009.
- ↑ Dityatev Alexander. Molecular Mechanisms of Synaptogenesis (англ.) / El-Husseini, Alaa. — New York: Springer, 2006. — P. 72—75.
- ↑ 1 2 3 4 Kinoshita T., Fujita M. Biosynthesis of GPI-anchored proteins: special emphasis on GPI lipid remodeling. (англ.) // Journal of lipid research. — 2016. — Vol. 57, no. 1. — P. 6—24. — doi:10.1194/jlr.R063313. — PMID 26563290.
- ↑ 1 2 Ikezawa Hiroh. Glycosylphosphatidylinositol (GPI)-Anchored Proteins // Biological & Pharmaceutical Bulletin. — 2002. — Т. 25, № 4. — С. 409—417. — ISSN 0918-6158. — doi:10.1248/bpb.25.409.
- ↑ Kinoshita T. Biosynthesis and deficiencies of glycosylphosphatidylinositol. (англ.) // Proceedings of the Japan Academy. Series B, Physical and biological sciences. — 2014. — Vol. 90, no. 4. — P. 130—143. — PMID 24727937.
- ↑ Coonrod S. A., Naaby-Hansen S., Shetty J., Shibahara H., Chen M., White J. M., Herr J. C. Treatment of mouse oocytes with PI-PLC releases 70-kDa (pI 5) and 35- to 45-kDa (pI 5.5) protein clusters from the egg surface and inhibits sperm-oolemma binding and fusion. (англ.) // Developmental biology. — 1999. — Vol. 207, no. 2. — P. 334—349. — doi:10.1006/dbio.1998.9161. — PMID 10068467.
Ссылки
[править | править код]- Чугунов, Антон; Полянский, Антон. Липидный фундамент жизни. Biomolecula.ru (22 января 2012). Дата обращения: 21 марта 2018.
Структуры клеточной мембраны | |
|---|---|
| Мембранные липиды[англ.] | |
| Мембранные белки | |
| Другое | |
Эта статья входит в число добротных статей русскоязычного раздела Википедии. |